آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Division Progymnospermophyta |


|
|
Read More
Date: 24-11-2016
Date: 23-11-2016
Date: 24-11-2016
|
Division Progymnospermophyta
PROGYMNOSPERMS
trimerophytes were an important group of plants that evolved from rhyniophytes . One evolutionary trend initiated in trimerophytes was pseudomonopodial branching, in which one branch dominates the plant as a trunk and other branches are smaller and act as lateral branches. Branch orientation was controlled such that regular spiral, decussate, and distichous branch systems resulted, various species of the genus Pertica being excellent examples. As that terminal branches became webbed, the first stages of megaphyll evolution were established. These evolutionary trends were continued in horsetails (division Arthrophyta) and ferns (division Pferidophyta), which evolved out of trimerophytes.
A third group to evolve from trimerophytes was the now-extinct progymnosperms, so named because some gave rise later to conifers, cycads, and the other gymnosperms (Table). Like ferns and horsetails, progymnosperms also developed megaphyllous leaves. However, another feature was just as significant—the evolution of a vascular cambium with unlimited growth potential and capable of producing both xylem and phloem.
Vascular cambia had also arisen in lycophytes and arthrophytes, but in both cases those cambial cells could not undergo radial longitudinal division. As wood accumulated, cambial cells were stretched tangentially and finally stopped functioning while the stem was still relatively narrow, less than 50 cm in diameter. Also, the vascular cambium produced either little or no secondary phloem.
The vascular cambium that evolved in progymnosperms was different. As early as 360 million years ago it was capable of undergoing radial longitudinal divisions, so it could [unction indefinitely, producing large amounts of both secondary xylem and phloem (Figs. 1 and 2). The wood was almost indistinguishable from that of many living conifers. It contained elongate tracheids, most with circular bordered pits. It had little or no axial parenchyma. Rays were tall and uniseriate and consisted of procumbent ray tracheids. This wood must have been strong, efficient at conduction, and capable of supporting a large mass of leaves and branches. At least some species, Triloboxylon and Proteokalon for exam pie, had a cork cambium that produced bark. Progymnosperms produced true woody trees: Trunks were up to 1.5 m in diameter and 12 m tall.

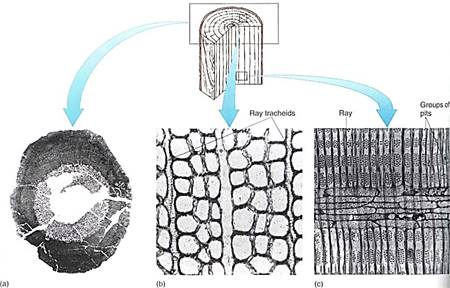
FIGURE 1: (a) Unlike the small amounts of wood produced in arborescent lycophytes (see Fig. 23.20a) and arthrophytes (see Fig. 23.29b), the vascular cambium of progymnosperms was able to produce a virtually unlimited amount of both secondary xylem and secondary phloem, as in this Callixylon brownii. (b and c) Progymnosperm wood had large vertical tracheids with pitted walls and an abundance of rays with ray tracheids. Ray tracheids do not occur in flowering plants but are common in conifers as horizontal tracheids that permit rapid conduction horizontally in the sapwood. (b) Callixylon erianum(X 700). (c) C. newberryi(X 375). (Courtesy of C. B. Beck., University of Michigan)

FIGURE 2: Reconstruction of Archaeopteris, a small tree about 6 m tall. Although extinct now, Archaeopteris flourished about 360 million years ago across northern North America, Canada, and Europe.
Although Progymnosperm wood was similar to that of gymnosperms, the two groups must be kept separate because progymnosperms did not have seeds or even ovule precursors. The species that arose later were heterosporous, but megaspores were shed through longitudinal slits in the sporangia; they were not retained in an indehiscent sporangium as occurs in seed plants. Although leaves and wood of progymnosperms were quite advanced, their reproduction was remarkably simple.
ANEUROPHYTALES
The order Aneurophytales contains the more relictual progymnosperms, such as Aneurophyton, Protopteridium, Proteokalon, Tetraxylopteris, Triloboxylon, and Eospermatoptem (Fig. 3). They varied in, stature from shrubs (Protopteridium, Tetraxylopteris) to large trees, up to 12 m tall. They all had a vascular cambium and secondary growth, but the primary xylem of the stems was a protostele, that is, a solid mass of xylem without pith; rhyniophytes and trimerophytes also had protosteles. The aneurophytes further resembled trimerophytes in having little webbing between their ultimate branches—telomes—and only in Proteokalon were telomes arranged in a plane. These could not yet be called leaves. These species were homosporous, all spores being produced in sporangia located at branch tips.
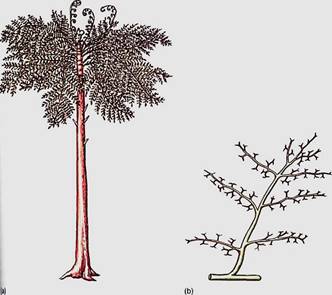
FIGURE 3: (a) Eospermatopteris was a member of the Aneurophytales; it had a trunk and what appear to be frond-like leaves, but these were just branch systems; planation and webbing had not yet evolved. (b) A portion of the "leaf1 of Aneurophyton. Planation had begun, but no webbing or sheet of photosynthetic tissue had yet appeared.
ARCHAEOPTERIDALES
A more derived progymnosperm was Archaeopteris in the order Archaeopteridales. These were trees up to 8.4 m tall with abundant wood and secondary phloem (2). Unlike aneurophyte stems, the stems of Archaeopteris had a siphonostele, pith surrounded by a ring of primary xylem bundles, much like modern conifers and dicots. Although the "fronds" of archaeopterids resembled fern fronds, close examination reveals that they were actually planated branch systems and that the ultimate "pinnules" were really spirally arranged simple leaves. Webbing is only partial in A. macilenta (Fig. 4), but it is complete in A. halliana and A. obtusa; these can be considered full-fledged megaphylls. Whether the leaves were annual or perennial is not known; the branches that bore the leaves did ret undergo secondary growth but rather remained herbaceous.
Reproduction in archaeopterids was still simple, but it was heterosporous. Megaspores measured up to 300 jam in diameter, microspores only 30 fim in diameter, and each type was produced in its own distinctive sporangium—broad megasporangia and slender microsporangia. The sporangia were terminal on short branches that were mixed with sterile, leaf-like branch systems. Megaspores were released from the sporangia, not retained. Seeds were not produced.
Within the progymnosperms the fundamental characters of stems, leaves, and secondary tissues had been established. The current hypothesis is that the Archaeopteridales later gave rise to the ancestors of modern conifers, and the Aneurophytales produced the ancestors of cycads and related plants .

FIGURE 4:During the existence of the plants classified in the extinct genus Archaeopteris, the final transitions from telomes to leaves occurred. (a) Archaeopteris fissilis. (b) A. macilenta. (c) A. halliana. (d) A. obtusa. What appear to be veins in the leaves were actually the shoot axes of the telomes.
EVOLUTION OF SEEDS
Investigating the life cycle of extinct plants based on fossil material is difficult, but not impossible. In freesporing species, spores can be identified with sporophytes only if some spores are trapped in a sporangium that was attached to leaves or wood during fossilization. Spores cannot be identified with gametophytes except when the gametophyte is microscopic and develops within the spore wall. If megaspores are retained in the megasporangium, at least some of the investigation is easier.
At present, the earliest known progymnosperm species with heterospory is Chauleria from the Middle Devonian Period, about 390 million years ago. It appears to have been a member of the Aneurophytales, and its sporangia contained both small (30 to 40 µm) and large (60 to 156 µm) spores. The slightly more recent Archaeopteridales (Upper Devonian Period, 360 million years ago) were heterosporous and had distinct microsporangia and megasporangia. In Archaeopteris halliana the number of megaspores in each megasporangum was low. Another Upper Devonian fossil, Archaeosperma arnoldii, was more derived: The megasporangium produced only one megaspore mother cell, and this produced only one large, viable megaspore and three small, aborted cells. The megasporangium was surrounded by a layer of tissue, an integument, that projected upward. There was a large micropyle, a hole in the integument that permitted the sperm cells to swim to the egg after the megaspore had developed into a megagametophyte and had produced eggs (Fig. 5).

FIGURE 5:A reconstruction of the megasporangium (b) and adjacent telomes (a) of Archaeosperma arnoldii. Each sporangium contained only one large megaspore; the other three products of the spore mother cell meiosis apparently degenerated early. Tissue immediately around the megaspore was the megasporangium wall. Attached to it and extending upward as finger-like projections was a layer called an integument. The exact nature of the integument is not known, but it may have been derived from telomes adjacent to the sporangium (see Fig. 6). Above the sporangium was a space surrounded by integument tissue, a calm, wind-free pollen chamber where pollen or spores settled. Around each megasporangium and integument was another set of partially fused telomes.
In Archaeosperma the integument was fused to the sporangium and was solid except at the tip of the micropyle, where it was lobed. In this condition it is difficult to determine how the integument arose evolutionarily, but in other fossils, early stages are present. In Genomosperma kidstoni the megasporangium was closely surrounded by sterile telomes (Fig 6). In G. latens the telomes were fused at the base, and in Eurystoma angulare they were similar to Archaeosperma, fused into one structure except at the tip. The sheath of sterile branches must have been important in trapping windblown microspores, but at first it would have allowed them to settle anywhere on the megasporangium. As they fused to each other and to the megasporangium, the space at the top of the megasporangium became the place where microspores settled, acting as a pollen chamber or holding area.
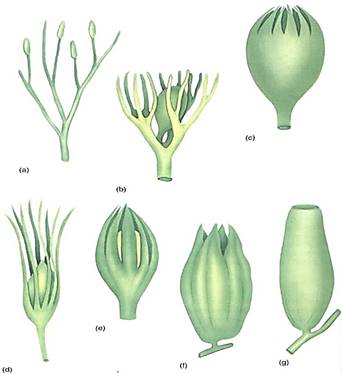
FIGURE 6 :(a to c) A hypothetical evolution of an integument from telomes. In (a), sporangia are neither clustered nor particularly associated with sterile telomes. (b) One megasporangium is surrounded by sterile telomes that all retain their individuality. (c) The sterile telomes have fused at their bases, forming a protective sheath around the megasporangium. (d to g). Actual fossils that correspond to the hypothesis. (d) Genomosperma kidstoni. (e) G. latens. (f) Eurystoma angulare. (g) Stamnostoma hyttonense.
As microspores evolved into pollen grains, changes occurred in the nature of their wall, in their method of germination, and in the nature of the microgametophyte they produced. By the Carboniferous Period, just after the Upper Devonian Period (about 340 million years ago), four types of pollen had become common. In some, internal structure is preserved well enough to see that these grains already had internal microgametophytes remarkably similar to those of modern gymnosperms.
As the megasporangia evolved into ovules with integuments, nearby telomes, either on the branches with the sporangia or on adjacent sterile branches, became modified into cupules (see Fig. 24.7a). These tended to be leaf-like and to surround the ovules loosely, perhaps giving some protection and helping create wind patterns that would bring in pollen grains. The cupules may have later given rise to the carpel in angiosperms.



|
|
|
|
دخلت غرفة فنسيت ماذا تريد من داخلها.. خبير يفسر الحالة
|
|
|
|
|
|
|
ثورة طبية.. ابتكار أصغر جهاز لتنظيم ضربات القلب في العالم
|
|
|
|
|
|
|
سماحة السيد الصافي يؤكد ضرورة تعريف المجتمعات بأهمية مبادئ أهل البيت (عليهم السلام) في إيجاد حلول للمشاكل الاجتماعية
|
|
|