























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Introduction to Seed Plants II: Angiosperms
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 23-11-2016
23-11-2016
 2892
2892




+
-
20
Introduction to Seed Plants II: Angiosperms
Flowering plants, the most recently evolved and most derived of all plant groups, are classified together in a single division, division Magnoliophyta, also known as division Anthophyta. The earliest fossils clearly recognizable as parts of flowering plants are pre-served pollen grains in rocks more than 130 million years old. We have not found any fossilized leaves or flowers from that time, so we believe the earliest flowering plants must have lived in drier areas where few sites were suitable for fossilizing plant parts, such as large, deep lakes full of fine-grained sediments. The first angiosperms probably were small trees or shrubs, because plants of this type survive well in drier regions and because the most likely ancestors—other seed plants in the gymnosperm groups—are woody trees.
The evolutionary transformation of a gymnosperm into a flowering plant was not a simple process, and it involved many alterations. The most obvious, of course, was the conversion of gymnospermous sporophylls into stamens and carpels, resulting in the formation of flowers. Because gymnosperms have rather flat, leaf-like sporophylls, those flowering plants that have similar reproductive parts are believed to be the most relictual. For example, groups such as Magnolia, Austrobaileya, and Degenariahave flat stamens without distinct filament and anther portions, and the sporogenous tissues, the microsporocytes (microspore mother cells), form relatively large, prominent internal masses (Fig. 1).
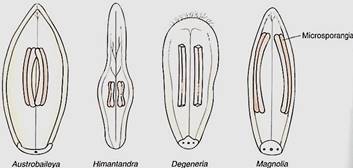
FIGURE 1:Several groups of angiosperms have stamens with what are believed to be relictual features; these features have not undergone much evolutionary change, so they are still similar to the ancestral features. The stamens (microsporophylls) of these four living dicots are much fleshier, flatter, and more leaflike than most other stamens of living species. Also, the sporogenous tissue is rather massive, similar to that of cycads and cycadeoids. Austrobaileya, Himantandra, and Degeneria are unfamiliar to most students; they are plants of South Pacific islands.
Carpels in these three genera are similarly leaflike, resembling young leaves whose blades have not yet opened. Instead of a stigma and style, the ovary edges have rows of hairs that function as a stigmatic surface (Fig. 2). Whereas ovules in gymnosperms are exposed on the megasporophyll surface, the folded nature of the sporophyll in flowering plants offers the ovules some protection; the carpels are closed or angiospermous, as opposed to the gymnosperm's open sporophyll. Although botanists often speak of the angiospermous carpel protecting the ovules, probably much more important is the fact that the pollen tube (microgametophyte) must now interact with the stigmatic area in addition to the nucellus, allowing more opportunity for the sporophyte to interact with the pollen and "test" it. Weak pollen or that of the wrong species can be inhibited from germinating, and the sporophyte can permit only healthy pollen of the proper species to pass through the stigma and reach its ovules (Fig. 3). Fewer ovules are wasted by exposure to improper or unhealthy pollen.
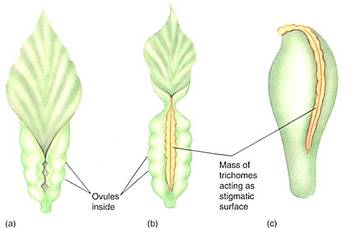
FIGURE 2:Carpels are megasporophylls, so we expect the most relictual living carpels to be rather leaflike, somewhat resembling megasporophylls of cycads . Several living angiosperms do have carpels resembling young leaves that have failed to open (a); the two halves of the blade are pressed together, a rather large amount of vascular tissue is present, and a stigma and style are absent. Hairs along the blade margin act like a stigma. If such a carpel produced ovules only at the base and if the upper part elongated, it would begin to resemble a more derived, modern carpel (c).
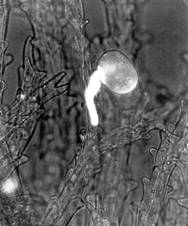
FIGURE 3:In many species of angiosperms, complex compatibility interactions occur between either the pollen and stigma or the pollen tube and stigma/style. In some cases, if the stigma and style detect an improper pollen tube, they deposit callose around it and inhibit its growth. The callose block is visible here as a white sheath (X 60). (Courtesy of H. L. Mogensen, Northern Arizona University).
In the transition from gymnosperms to angiosperms, fertilization evolved such that the second sperm cell of the pollen tube fuses with the polar nuclei of the megagametophyte, producing the endosperm nucleus. This process of double fertilization is universal in flowering plants. Within the vegetative body, the major transitions were the evolution og vessel elements and sieve tubes (Fig. 4). Leaves became broader and developed reticulate venation, and they became more polymorphic, adaptable to a variety of functions, not just photosynthesis.

FIGURE 4: (a) In living plants, vessel elements that most closely resemble tracheids are long and narrow; short, wide vessel elements are derived. Most long, narrow vessel elements have scalariform perforation plates, so we assume that these vessel elements evolved from tracheids with scalariform pits. (b) Because a sieve element conducts mostly longitudinally, it is most efficient if the sieve areas and sieve pores on its ends are especially large. When this finally occurred, sieve tube members came into being.
Structural modifications are most easily seen and studied, but probably a more fundamental transition was the acquisition of developmental plasticity, the capacity to survive mutations that alter growth and development. Conifers lack developmental plasticity; £r example, most people never have trouble recognizing an unfamiliar plant as being a conifer when they see one because all conifers greatly resemble each other. They are all large or giant trees with one or a few main stems and needle- or scale-shaped leaves. They are always green or brown, never brightly colored except for the red seeds of yew and Ute gray-blue berry-like cones of junipers. This uniformity is even more striking when one considers that gymnosperms dominated Earth for hundreds of millions of years and could have diversified into almost any habitat. On the other hand, angiosperms have tremendous developmental plasticity: Beginning about 125 to 145 million years ago, they have diversified so greatly that not only hundreds of thousands of species, but also thousands of types, now exist. They range from gigantic (Eucalyptus, oaks, elms) to tiny (mustards, certain daisies; Fig. 5a); woody to herbaceous; perennial to biennial to annual; temperate to tropical; desert habitats to mesic to rainforest to aquatic; autotrophic to partially parasitic to fully parasitic; and epiphytic to subterranean (Fig. 5b) to endophytic (living inside another plant, such as Rafflesia, Tristerix).

FIGURE 5: (a) This dwarf daisy (Monoptilon) is by no means the smallest flowering plant. Many species of mustard, often the first flowers of spring, are also tiny. (b and c) Ombrophytum subterraneum from the town of Chiu Chiu in northern Chile, the southernmost outpost of the Inca empire. Ombrophytum is a parasite on roots of surrounding plants and is reported by people of the area never to emerge above ground. The plant in (c) consists of an irregular, lumpy "tuber" and two inflorescences with tiny pink flowers. The root and stem base of the host are visible. (Courtesy of Eliana Belmonte, Universidad de Tarapaca).
The evolutionary changes involved in the conversion of a gymnosperm line into an angiosperm line did not occur instantaneously, nor did they occur in all species. Double fertilization and the ability to form flowers were probably the first transformations, because all flowering plants have these features. Sieve tubes possibly evolved next because only one or two species are thought to lack them. But the flowering plants must have existed and diversified for several million years before vessel elements evolved, because several families do not have vessels; these families must have evolved and become distinct before vessels appeared. All the herbaceous species today do have vessels, so herbaceousness must have developed next, and it probably arose several times in many separate lines. The ability to produce flowers in the first year after seed germination is a necessary trait for annual plants, most of which are herbs. Only much later did characteristics such as succulence, parasitism, and vining shoots develop; only a few isolated families or species have these characters.
Other derived features are the fusion of the petals into one structure (sympetally) and floral zygomorphy, that is, flowers that are bilaterally symmetrical, not radially symmetrical (Table). By this time the flowering plants had diversified so much that each of these evolutionary innovations involved only a small part of the whole group. It should not be thought that the flowering plants are now "finished," that all their evolution has already happened. Some groups do seem to be changing very little now, but others, especially grasses, composites, bromeliads, and orchids, are still changing and evolving so rapidly that it is difficult to keep up with them. Similarly, we have no reason to expect that the flowering plants will always be the most derived group; lycophytes dominated Earth for millions of years, only to be displaced by gymnospermous seed plants, which in turn were overshadowed by flowering plants.
A special word of caution is necessary about the flowering plants. Angiosperms have evolved in numerous directions and undergone many changes. For example, the original condition of early angiosperms is believed to have been large shrubs or small trees; some evolutionary lines appear to have retained this feature, so the shrubby plants of those lines are relictually shrubby. But other lines evolved to be herbaceous or nearly so (a derived feature), but then some of the descendants of these herbs became woody and shrubby again. Although they may appear homologous to the relictually shrubby plants, in these lines scrubbiness is actually a derived feature—they are secondarily shrubby. Because of the large amount of evolutionary change that has occurred in the angiosperms, we must always be careful not to mistake analogous features for homologous features.

 الاكثر قراءة في البذور (مغطاة البذور - عاريات البذور)
الاكثر قراءة في البذور (مغطاة البذور - عاريات البذور)
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















