آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| انظمـــــة اصلاح DNA |


|
|
أقرأ أيضاً
التاريخ: 2025-02-18
التاريخ: 2025-02-18
التاريخ: 2025-02-02
التاريخ: 10-1-2016
|
انظمـــــة اصلاح DNA
التغيرات التي تحصل في المواد الوراثية قد تؤثر في أكثر من جانب، فهي قد تكون مميتة للخلايا او تؤدي الى تغيرات ليست في صالح الخلية، ولكن قد تكون في صالح الاستعمال لانتاج المواد، ومن جهة اخرى فهي تمثل رافدا مهما في عمليات تطوير الأنواع، وفي الاحياء المجهرية وخاصة الصغيرة مثل البكتريا التي تمثل أبسط وأصغر الاحياء يجب ان تكون لها أنظمة اصلاح لمادتها الوراثية كفوء جدا وذلك لان الخلايا بدائية النواة تمتلك جينوم فرادني وبالتالي نسخة واحدة من كل جين. وأغلب مواده او شفراته فعالة، ولذلك فأي ضرر يصيب الجينوم يكون ذات نتائج كارثية على الخلايا ان لم تكن مميتة، ولذا يظهر على الاحياء التطبع لتستمر بالحياة.
وهناك العديد من العوامل البيئية يمكن ان تؤثر على الخلايا، لعل أهمها هو دخول الخلايا الى طور الاستقرار حيث تزداد الاعداد والفضلات وهذا يؤدي الى ظهور الطفرات، اضافة الى عوامل بيئية اخرى يمكن ذكر بعضها بشكل مبسط .؟
• الأشعة.
• الجفاف.
• التعرض للمواد الكيماوية المؤذية.
• الحرارة.
• المضادات الحيوية المستعملة في العلاج او المنتجة في البيئة.
كلها يؤدي حدوث الاضرار في DNA وبحالات مختلفة وبعض الأحيان تنتج :
1- كسور في واحد أشرطة DNA وهذه يمكن تلافيها بتأثير الانزيمات اللاحمة وعادة لا تؤدي الى موت الخلايا.
2- احداث كسور في أشرطة DNA المزدوجة وتكون مميتة في أغلب الأحيان ولا يمكن ربطها بالأنزيمات اللاحمة لذلك تفكك بالأنزيمات المحللة للمواد الوراثية Nucleases.
3- احداث تغير في القواعد ينتج عن ضرر تأكسدي والذي ينتج أيضاً من فعالية الخلايا نفسها مثل عمليات التنفس في الاحياء الهوائية وعادة تكون مميتة لأنها تكون بمثابة حواجز امام عملية تضاعف الاشرطة عند نقطة الضرر.
لذا فالضرر يتخذ اشكالات مختلفة، لذلك وجدت انظمة اصلاح مختلفة اضافة الى ان تعدد الانظمة يكون افضل من وجود نظام اصلاح واحد، وتتسم معظم انظمة الإصلاح بصفات مشتركة وهي التعرف على الضرر او الكشف عنه، ثم اجراء الاصلاحات اللازمة ويمكن ان يعمل أكثر من نظام الصلاح في التخلص من الضرر الحاصل فمثلا الضرر الحاصل من الاشعة فوق البنفسجية تؤدي الى ارتباطات في الشريط الواحد بتكوين تراكيب حلقية بين البريميدنيات وخاصة الثايمين تؤدي الــــــــــــــــــى اضطراب فـــــــــــي حلزون DNA وتمنع انزيم كوثرة (DNA polymerase) DNAP III من اجراء عملية التضاعف عندما يصل الى منطقة الضرر ويتوقف، ولكن التوقف يكون آني نظراً لوجود علميات الإصلاح ومنها :
1- الإصلاح الضوئي Photoreactivation : نظام اصلاح مباشر وفيه يقوم الجين phr بالتشفير للأنزيم الخاص Deoxyribodipyrimidine photolyase وبمساعدة حامض الفوليك كمتمم انزيمي يقوم بالارتباط الى مزدوج الثايمين في الظلام وعند وجود الضوء يمتص حامض الفوليك الضوء المرئي ويستعمل الطاقة لكسر الأواصر بين قواعد الثايمين المرتبطة ثم يترك الانزيم DNA.
2- الإصلاح الاستئصالي Excision repair : نظام اصلاح مباشر ويشارك النظام في اصلاح الضرر الناتج من عدة عوامل ويسمى النظام بإصلاح ضرر الاشعة فوق البنفسجية بـ (NER) Nucleotide excision repair وفيه يقوم أحد الانزيمات القاطعة الداخلية والذي يسمى Correndonuclease (انزيم القطع الداخلي المصحح) والذي يمكن ان يميز مزدوجات الثايمين ويثلم الشريط عند النهاية 5'. ويتكون الانزيم من عدة وحدات ثانوية هي منتجات الجينات uvrA، uvrB، uvrC، uvrD، والبروتينات الناتجة يتداخل بعضها ليكون جسيم الإصلاح Repairosome.
يرتبط البروتين uvrA مع ATP الى DNA عند المنطقة الملوية او المشوهة، ثم يقوم البروتين uvrB بالارتباط للمعقد المتكون والذي يزيد من تخصص المعقد uvrA-ATP للـ DNA المشعع. اما البروتين uvrC فيقوم بقطع شريط DNA في منطقة تبعد حوالي 8 قواعد على يسار (Upstream) منطقة المزدوج البريمديني وكذلك يقطع الشريط على بعد 4-5 قواعد من جهة اليمين (Downstream) . اما بروتين uvrD والذي هو DNA helicase Π (والذي يشابه Dna B المستعمل عند بدء التضاعف) فيقوم بفصل الاشرطة وفك تركيبها الحلزوني لأطلاق القطعة التي تتكون من حوالي 12 – 13 زوج قاعدة. ثم يقوم بعد ذلك DNAPI وغيره من الانزيمات بمليء الفجوة بالاتجاه 5' الى 3' ثم لحمها. وفي حالة عدم الإصلاح فان DNAPIII يبدأ بمضاعفة اشرطة DNA بعد منطقة الضرر تاركا منطقة مصابة التي قد تصل الى حوالي 800 زوج من القواعد ويطلق عليها عبور الاصابة Translesion وعندها تغطى منطقة الاصابة او الفجوة ببروتينات RecA التي تغطي اشرطة DNA المفردة الى حين قيام انظمة اخرى مثل اصلاح التأشب Recombination repair والتي تتم بعد التضاعف لذلك تكون الطفرات الخالية من الجين recA حساسة جداً للأشعة فوق البنفسجية وهذا التصحيح او الإصلاح هو جزء من نظام SOS.
ومن أنظمة الإصلاح الاخرى ومنها :
• Ap repair system : ويقوم بإجراء الإصلاح على DNA الذي ازيلت منه قاعدة بيورينية بواسطة الانزيمات، والانزيمات العاملة في هذا النظام تسمى Ap endonucleases، والصنف الأول منها يثلم DNA عند النهاية 3' للموقع الذي فيه ضرر، والصنف الثاني يفلق الطرف 5'. وتزيل قطعة صغيرة من DNA ثم يقوم الانزيم DNAPI بمليء الفجوة والانزيم اللاحم Ligase يلحم القطعة الجديدة مع باقي الشريط. وهناك انزيمات متخصصة فمثلا Uracil-DNA glycosylase يقوم بإزالة قاعدة اليوراسيل من DNA وذلك بفلق الاصرة بين السكر والنتروجين لإزالة القاعدة.
• نظام اصلاح عدم التلاؤم (MMR) Mismatch repair system : ويكون النظام مسئولا عن عدد كبير من عمليات الإصلاح التي تعقب التضاعف، ويساهم بشكل كبير في عمليات التطور ويتكون النظام من عدة جينات mut S و mut L و mut H و mutU التي تشارك نواتجها في عمليات الإصلاح. Mut اشتقت من mutator لأنه عند حصول طفرات في جينات النظام فان الخلايا تعاني من زيادة في مستوى الطفرات التلقائية. ويلعب النظام دورا هاما في حياة الخلايا والاحياء التي يوجد فيها والتي تمتد من البكتريا الى الاحياء الراقية مثل اللبائن. فهو يساعد في العديد من العمليات منها :
1- تجنب حصول الطفرات في الخلايا.
2- يساعد في اعادة ترتيب الجينوم.
3- ينظم انتقال الجينات الافقي بين الانواع.
4- يساعد في منع تطور السرطانات في الاحياء الراقية.
وتتم فعالياته بعدة خطوت، تبدأ بتمييز والتعرف على المناطق التي أدخلت فيها القاعدة غير الصحيحة ثم استئصالها واستبدالها بواحدة صحيحة. وعملية التمييز تعتمد على مثيلة اشرطة DNA الجديدة التي يقوم بها الانزيم DNA methylase الذي يشفر له بالجين dam، والانزيم يقوم بمثيلة التوالي 5'-GATC-3' عند القاعدة الادنين، ولذلك تقوم منتجات الجينات mut L، mut S بالتعرف على حدوث الخطأ. اما ناتج الجين mutU وهو DNA helicase II (الذي يسمى اضا جين uvr D) بفك التواء اشرطة DNA المزدوجة مطلقا المنطقة المثلومة ثم تملأ الفجوة بفعالية DNAPI والانزيمات اللاحمة.
نظام SOS :
او ما يسمى بنظام الاستغاثة (Save Our Souls) ويكون بمثابة منظم كبير SOS regulon يستجيب للضرر الذي يحصل في DNA وتعد الاستجابة من النوع العام Global transcriptional response وتؤدي الى حث ما يقرب من 20 جين تعمل في مسارات مختلفة من مسارات اصلاح DNA والموضحة في الشكل (1).
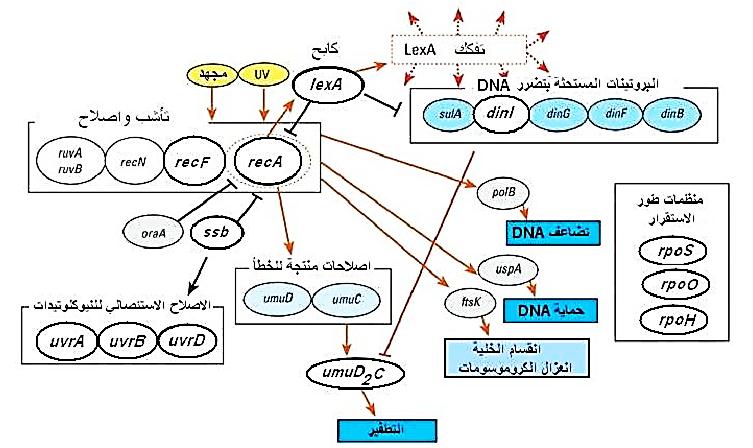
شكل 1 : نظام الاستغاثة SOS
وتحصل استجابة الاستغاثة عندما يحصل ضرر DNA يفوق قابلية الخلايا للإصلاح بواسطة الانظمة الاخرى وعند تجمع DNA غير المصحح في الخلايا. وتكون الاستجابة مرافقة الى حث مجموعة من الاستجابات الفسلجية. ويساعد النظام الذي درس في E.coli بشكل مفصل في حصول عمليات اصلاح التأشب وكذلك الإصلاح الاستئصالي وفي الوقت نفسه يشجع ظهور الطفرات نظراً لكون عمليات الإصلاح من نوع المرافقة للأخطاء، فمثلا عند التعرض للأشعة فوق البنفسجية وحدوث عمليات الإصلاح الضوئي او الاستئصالي او بعد التضاعف فمعظمها لا تؤدي الى حدوث الطفرات ولكن اشتراك نظام الاستغاثة يساعد على حدوث الطفرات ويكون مرافقا لتوقف الانقسام مؤديا الى انتاج خلايا بشكل خيوط.
ويوجد منتجان من جينات النظام وهما بروتين التأشب RecA يعمل حاثا للنظام وبروتين التأشب له وظائف متعددة اضافة الى عمله في تصحيح DNA فهو يقوم بقدح او بدأ الفعاليات المحللة للبروتينات الداخلية في الخلية. والبروتين الاخر هو LexA الذي يعمل كابحا للنظام، والجينات في النظام تحوي على SOS box يرتبط به البروتين الكابح والبروتين الاخير يكون من البروتينات ذات التنظيم الذاتي. والاحداث المحفزة للنظام هي وجود اشرطة مفردة من DNA، ويكون التحسس ببروتين التأشب مؤثراً على الكابح الذي يعاني من التحلل الذاتي ويصبح غير قادرا على الارتباط بالجينات العاملة في النظام ويصبح بالإمكان قراءتها وانتاج البروتينات. ويقوم النظام بتصحيح DNA المتضرر بعدة طرق :
1- يقوم النظام بتغطية المنطقة المتضررة اثناء عمليات التضاعف، ثم بعد ذلك يتم التعامل مع هذه المنطقة في الاشرطة الأبوية بواسطة الإصلاح التأشبي والاصلاح بعد التضاعف Post replicative repair.
2- عملية اصلاح تتجاهل قاعدة ازدواج القواعد المعروفة في اشرطة DNA وهذه تسمى التصحيح المولد للأخطاء (Translesion DNA synthesis) Error –porne repair وقد تؤدي الى حصول على شفرات غير حساسة Non sense codon وانتاج بروتين غير فعال نتيجة لوجود حامض او حوامض امينية بشكل خاطئ. ويحوي النظام على جينات اخرى كما موضح في الشكل، هي sulB و SulA التي تنشط وتعمل على منع انقسام الخلايا لإعطاء وقت كافي لإصلاح الضرر في DNA قبل بدء التضاعف. وحدوث الطفرات بعد عمل نظام الاستغاثة يعود الى سرعة عمليات التضاعف وقلة قابلية وفعالية Proof reading، ولكن مع هذا يلاحظ ان الخلايا التي لا تحوي على نظام الاستغاثة او كونه معطلا بالطفرات فان الخلايا تموت عند تعرضها للعوامل المدمرة لموادها الوراثية.
واثناء عمل نظام الاستغاثة يلاحظ ان بروتين التأشب يمكن ان تؤثر على كابح العاثيات الاولية المرتبطة بكروموسومات الخلايا مؤديا ذلك الى إلغاء حالة الاستذابة ودخول العاثي دورة التحلل، وبعد اجراء عمليات التصليح فان بروتين التأشب لا يقوم بحيث البروتين الكابح الذي لا يتحلل ويتجمع في الخلية ليعود لغلق الجينات العاملة في نظام الاستغاثة. وهناك عدة عوامل تؤثر في حث النظام منها قلة المواد الغذائية او ارتفاع الضغط التنافذي حول الخلايا وبعضها يمكن ان يجعل الاستجابة سريعة وتصل اقصا حالاتها بعد حوالي 30 دقيقة. اضافة الى وجود ظروف اخرى منها معاملة الخلايا ببعض المضادات الحيوية والاجهاد الحراري ويتم ذلك في الخلايا التي تقوم بالتنفس الهوائي اذ يتوقف حثه تحت ظروف تنفس النترات ويعود حثه عند عودة الظروف الهوائية. ومن الملاحظ ان المواد السامة للمواد الوراثية في الاحياء حقيقية النواة الراقية تحث الاستجابة في البكتريا وهذه العلاقة هي الاساس في العديد من الفحوص لاختبار قابلية المواد للتطفير و التسرطن على غرار اختبار ايمس Ames test. وفي هذه الحالة يستخدم الاختبار اللوني SOS - chromo test.
واستجابة الاستغاثة في البكتريا الاخرى تكون متشابهة في الاساسيات لما ذكر اعلاه مع بعض الاختلافات فمثلا في B. subtilis فان النظام يؤدي الى حث ظاهرة التأهل Competence في الخلايا عند عدم وجود ضرر في DNA ويكون البروتين الكابح هو البروتين DinR ناتج الجين din ويرتبط بمنطقة المشغل للنظام SOS operator.
المصادر
الخفاجي , زهرة محمود (2008) . التقنية الحيوية الميكروبية (توجهات جزيئية ) . معهد الهندسة الوراثية والتقنية الحيوية . جامعة بغداد .



|
|
|
|
دخلت غرفة فنسيت ماذا تريد من داخلها.. خبير يفسر الحالة
|
|
|
|
|
|
|
ثورة طبية.. ابتكار أصغر جهاز لتنظيم ضربات القلب في العالم
|
|
|
|
|
|
|
سماحة السيد الصافي يؤكد ضرورة تعريف المجتمعات بأهمية مبادئ أهل البيت (عليهم السلام) في إيجاد حلول للمشاكل الاجتماعية
|
|
|