
آخر المواضيع المضافة

 الفيزياء الكلاسيكية
الكهربائية والمغناطيسية
علم البصريات
الفيزياء الحديثة
النظرية النسبية
الفيزياء النووية
فيزياء الحالة الصلبة
الليزر
علم الفلك
المجموعة الشمسية
الطاقة البديلة
الفيزياء والعلوم الأخرى
مواضيع عامة في الفيزياء
الفيزياء الكلاسيكية
الكهربائية والمغناطيسية
علم البصريات
الفيزياء الحديثة
النظرية النسبية
الفيزياء النووية
فيزياء الحالة الصلبة
الليزر
علم الفلك
المجموعة الشمسية
الطاقة البديلة
الفيزياء والعلوم الأخرى
مواضيع عامة في الفيزياء| دور ميكانيك الكم في عمل الإنزيم دي إن إيه بوليميريز |


|
|
أقرأ أيضاً
التاريخ: 2023-04-12
التاريخ: 2023-04-10
التاريخ: 2023-04-11
التاريخ: 2024-04-06
|
الإنزيم DNApolymerase
هو الإنزيم الذي يقوم بمضاعفة جزيئات الـ DNA الأصلية في الخلية قبل انقسامها؛ حيث تبقى نسخة من الجزيئة في الخلية الأم وتذهب نسخة مماثلة إلى الخلية الجديدة. (شكل 6-1) كما يقوم الإنزيم بتصحيح الأخطاء التي تحدث أثناء عملية تكوين النُّسَخ الجديدة. وبذلك تُحفظ المعلومات الوراثية المخزونة في جزيئات DNA بدرجة عالية من الدقة وتُنقل إلى الخلايا الوليدة. هذا يحصل في جميع الخلايا القادرة على الانقسام في الكائن الحي المتعدد الخلايا وكافة خلايا الكائنات الحية الوحيدة الخلية، وبذلك تتم المحافظة على النوع.
تحتوي جميع خلايا الأحياء البدائية النواة والأحياء الحقيقية النواة على بضعة أنواعٍ من إنزيمات DNApolymerase ، كما تحتوي جُسَيمات بعض الفايروسات على أنواع خاصة من هذه الإنزيمات تختلف هذه الإنزيمات في التركيب والوظيفة، لكنها مع ذلك تُحافظ على حقول تحفيز متشابهة تركيبيًا ووظيفيًّا (2001 ,Hingorani)؛ فبعضها يقوم بالبلمرة (المضاعفة) والبعض الآخر بالبلمرة والتصحيح من خلال امتلاكه لحقل إزالة النكليوتيدات (Exomuclease). في البكتيريا يُوجد ثلاثة أنواع من الإنزيم DNApolymerase هي pol I و pol Il و pol Ill ، الأخير هو الإنزيم الذي يقوم بالبلمرة الرئيسة. في الأحياء الحقيقية النواة ثمَّة 16 نوعًا من هذا الإنزيم، الرئيسة فيها هي α pol وδ pol وЄ pol.

شكل 6-1: تركيب الإنزيم DNA polymerase beta ثلاثي الأبعاد في الخلايا البشرية، مع توضيح ارتباط جزيئة DNA.
من أجل مضاعفة الـ DNA، يقوم الإنزيم DNApolymerase بإضافة النكليوتيدات إلى ذرة الكاربون 3 لشريط الـ DNA نكليويدة في كل خطوة، ما يؤدي إلى تكوين شريطي DNA أحدهما أصلي (يُستخدَم كقالب) والآخر جديد.
تبدأ العملية بفك شريطي الـ DNA المرتبطين بأواصر هيدروجينية بين القواعد النووية المتقابلة بواسطة الإنزيم Helicase ومساعدة الإنزيم Topoisomerase. كل شريط من شريطي الـ DNA الأصلية سيكون قالبًا لعمل الإنزيم. إن الإنزيم DNApolymerase يعمل بإضافة نكليويدة في كل خطوة إلى النهاية 3 من النكليوتيدة الأخيرة في الشريط الموجود، وبذلك سيسير باتجاه 3' ← 5' منتجًا شريطا جديدًا باتجاه 5' ← 3' موازيًا ومعاكسا لشريط القالب (شكل 6-2). الإنزيم DNApolymerase لا يتمكن من تكوين شريط DNA بدءًا من الصفر؛ لذلك لا بد من وجود شريط نكليوتيدات موجود أصلًا ليتمكن الإنزيم من الإضافة إليه. هذا الشريط هو البادئ (Primer) الذي يتألف من قواعد نووية لـ RNA أو DNA أو كليهما. القاعدتان النوويتان الأوليتان تكونان بشكل RNA وتُخلقان بواسطة الإنزيم Primase. النتيجة تكوين شريطين متممين لبعضهما؛ أحدهما أصلي والآخر جديد.
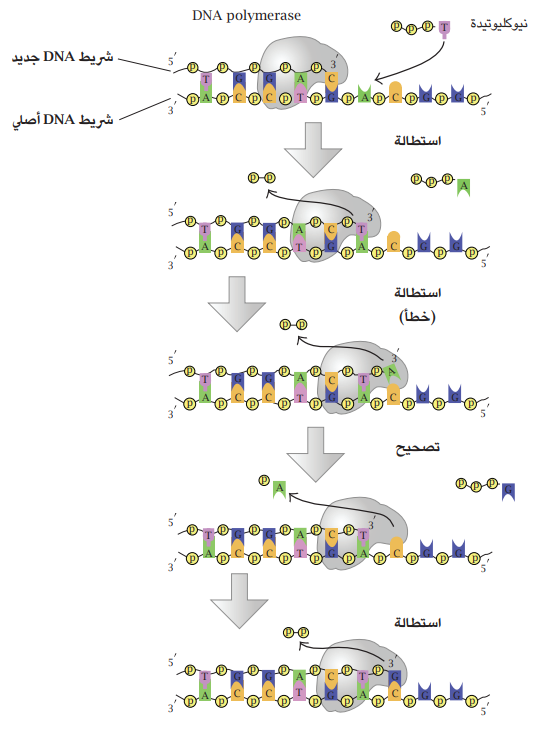
شكل 6-2: مخطط عمل الانزيم DNApolymerase في مد شريط DNA مع تصحيح الخطأ. عن I.Madprime ميكانيكا الكم وعمل الانزيم DNApolymerase.
إن دقة عمل الإنزيم DNApolymerase عالية كما سبق ذكره؛ فنسبة الخطأ؛ أي وضع قاعدة نووية خطأ (غير متممة لقاعدة القالب) هي بحدود واحد بالمليار. تقوم بعض أنواع هذه الإنزيمات بتصحيح الخطأ بواسطة حقل Endonuclease الذي يقوم بإزالة القاعدة النووية الخطأ ويُعاود الإنزيم استبدالها بالصحيحة. الخطأ هنا سيؤدي إلى إحداث طفرة؛ أي تغيير في تسلسل القواعد النووية في شريط الـ DNA الجديد، والتي يمكن أن يكون لها عواقب ضارة بحياتية الخلية غير أنها من ناحية أخرى تُسهم في إحداث التغاير الوراثي المطلوب من أجل التطور.
يعتمد هذا الجزء على عمل الباحثة (2008) Goel في دراستها لأهمية ميكانيك الكم في عمل إنزيمات البوليميريز. إنَّ إنزيمات DNApolymerase التي تُضاعف الـ DNA و RNApolymerase التي تُنسخ الـ DNA على شكل جزيئات RNA من أجل تكوين البروتينات وغيرها هي محركات نانوية (Nanomotor) كونها ذات أبعاد نانوية، وتتمكَّن من تحويل الطاقة الحرة إلى طاقة ميكانيكية. وهي معالجات معلومات أيضًا؛ حيث تقرأ المعلومات في جزيئة الـ DNA وتكتبها متأثرةً بالمعلومات التي في البيئة؛ كونها منظومات معقدةً متكيفية مفتوحة. البيئة هنا تشمل درجات الحرارة، وتركيز النكليوتيدات والشد الميكانيكي للجزيئة والتفافها والإشارات الكهرومغناطيسية ... إلخ، والتي يمكن أن تؤثر على ديناميكية هذه المحركات النانوية؛ فالتجارب تشير إلى أن شد جزيئة الـ DNA يمكن أن يؤثر على سرعة الإنزيم DNApolymerase ويمكن أن يعكس اتجاهه إذا زاد عن 35 بيكونيوتن (شكل 6-3).
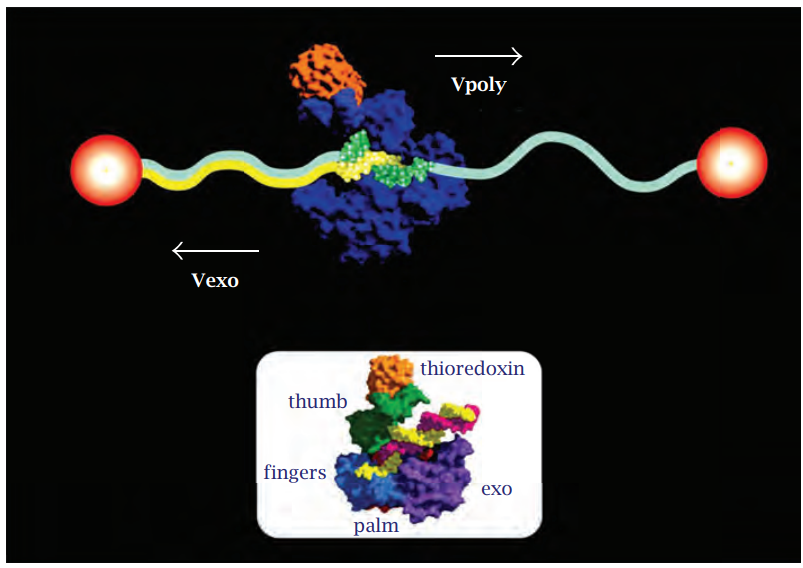
شكل 6-3: تشير تجارب الجزيئات المفردة أن الشد الميكانيكي فوق 35 بيكونيوتن على جزيئة الـ DNA يحفز سرعة عكسية في محرك إنزيم (T7 DNA polymerase (DNA للفايروس البكتيري. تم مطٌ شريط DNA مُفرَد بين خرزتين بلاستيكيتين في الوقت الذي يحفز الإنزيم تحويل الشريط المفرد SSDNA إلى شريط مزدوج dsDNA. أُشير إلى سرعة البلمرة أو تكوين dsDNA بـ Vpoly، بينما أُشير إلى القص العكسي لـ dsDNA بـ Vexo. الصورة السفلية هي التركيب البلوري لمعقد T7 والذي يُبيِّن أن نشاط محرك إنزيم البوليميريز والمحور يوكليز «مغير سرعة أمامي وخلفي ينتج عن حقلي تحفيز متميزين. عن: (2008) Goel.
التقدم التقني الذي مكَّن من دراسة ديناميكية هذه التراكيب على مستوى طول وزمن متزايد الصغر، يُمكّن من اختبار التأثيرات الكمومية تجريبيا، ولكي تتحقق التأثيرات الكمومية في عمل هذه الإنزيمات ينبغي أن يكون زمنُ فك التماسك (Decoherence) لها طويلًا وزمن التماسك (Coherence) قصيرًا كقاعدة عامة في تحقق الحالة الكمومية. باستخدام لامتساويات وغنر (1957 ,Wigner) الأولى والثانية التي توضّح المحدّدات التي يفرضها ميكانيك الكم على صحة ودقة المسافات والفترات الزمنية في الساعة الكمومية (المنظومة الكمومية) يمكن أن تُحدد أهلية أية ماكنة معالجة للمعلومات؛ فيمكن تحديد مدى صحة ((Accuracy المنظومة الكمومية زمن التشغيل الأقصى (Tmax)، ودقتها (Presision) الفترة الزمنية الصغرى (Tmin) التي يُمكن أن تُنجزها المنظومة الكمومية كدالة لكتلتها M وعدم التحديد في موقع الإنزيم على طول جزيئة الـ λ DNA وثابت بلانك ћ:

ويمكن استخدام لامتساويات وغنر لمعرفة الطول الأقصى Lmax الذي يتمكن الإنزيم من قراءته بشكل صحيح، وكذلك حجم الخطوة المؤثّرة الصغرى Lmin للإنزيم. لامتساوية وغنر الأولى تحدد المدى الزمني (والمدى الطولي) الذي يمكن فيه مضاعفة الـ DNA بشكلٍ صحيح مع بقاء المنظومة متماسكة كموميًا. أما لامتساوية وغنر الثانية فتضع حدودًا لدقة عمل الإنزيم وقدرته على معالجة المعلومات.
عند اتخاذ λ كطول جزيئة الـ DNA للبكتريوفاج لامبدا والبالغ 16مكم، فإن تطبيق لامتساوية وغذر الأولى سيكون Tmax للإنزيم بوليميريز هو أقل من 387 ثانية، وللمقارنة فإن DNApolymerase للبكتيريا (Thermus aquaticus (TAO هو خطأً واحد كل 100 ثانية. كما أن لا متساوية وغنر الثانية تُبيّن أن Tmin (أقل فترةٍ زمنية ممكن قياسها) هي 14–10 × 5 ثانية أو 500 فمتوثانية. إن Tmin لا يساوي زمن تضمين قاعدة نووية من قبل الإنزيم؛ وإنما هو زمن تحول حالة داخلية للإنزيم.
ويمكن الحصول على Lmax وهو أقصى طول من الـ DNA يمكن للإنزيم قراءته بشكل صحيح وتبين أنه 104 × 4 قاعدة نووية، وهو مقارب لما هو معروف عن الإنزيم TAQpolymerase والمساوي إلى خطأ واحد كل 104 قاعدة نووية يقرؤها. كما أن Lmin الذي يعني أقل مسافة يمكن فيها تسجيل معلومات بواسطة الإنزيم تساوي حوالي 12–10 × 5 قاعدة نووية، وتُعادِل 21–10 × 2م.
ومن خلال الحسابات ونتائج تجربة عمل الإنزيم على DNA البكتريوفاج لامبدا وطولها 16 مكم، فإن الوقت الأقصى اللازم لقراءة الـ DNA هو 500 ملي ثانية للشريط المزدوج، و3 ملي ثانية للشريط المفرد من الـ DNA بينما وقت فك التماسك الأقصى لمعقد الإنزيم – DNA يتراوح بين بضع دقائق إلى بضع ساعات. وهكذا يمكن الاستنتاج أن تأثيرات ميكانيك الكم تلعب دورًا مسبقًا في التأثير على ديناميكية قراءة الإنزيم للـ DNA. وتُظهر النتائج أيضًا أن كمية المعلومات أو عدد البتات المخزونة في منظومة الإنزيم –DNA ، هي أكبر بكثير مما هو مُتصوّر (باعتباره 1 بت / قاعدة نووية). وهذه الزيادة حسب الباحثة ترجع إلى أن الإنزيم يمتلك بضع حالات مجهرية داخلية. كما يُظهر هذا العمل أن المعلومات ليست مخزونةً في الـ DNA فقط، وإنما يُشكّل الـ DNA والإنزيم وبيئتهما شبكة معالجة معلومات ديناميكية معقدة ذات مخزون معلومات، وقدرات معالجة عالية جدا.



|
|
|
|
"عادة ليلية" قد تكون المفتاح للوقاية من الخرف
|
|
|
|
|
|
|
ممتص الصدمات: طريقة عمله وأهميته وأبرز علامات تلفه
|
|
|
|
|
|
|
تسليم.. مجلة أكاديمية رائدة في علوم اللغة العربية وآدابها
|
|
|
















