Coordinated Regulation of Glycolysis and Gluconeogenesis:- Glycogen Phosphorylase Is Regulated Allosterically and Hormonally
In the late 1930s, Carl and Gerty Cori (Box 15–1) dis covered that the glycogen phosphorylase of skeletal muscle exists in two interconvertible forms: glycogen phosphorylase a, which is catalytically active, and glycogen phosphorylase b, which is less active (Fig. 15–24). Subsequent studies by Earl Sutherland showed that phosphorylase b predominates in resting muscle, but during vigorous muscular activity the hormone epinephrine triggers phosphorylation of a specific Ser residue in phosphorylase b, converting it to its more active form, phosphorylase a. (Note that glycogen phosphorylase is often referred to simply as phosphorylase—so honored because it was the first phos phorylase to be discovered; the shortened name has persisted in common usage and in the literature.) The enzyme (phosphory lase b kinase) responsible for activating phosphorylase by transferring a phosphoryl group to its Ser residue is itself activated by epinephrine or glucagon through a series of steps shown in Figure 15–25. Sutherland discovered the second messenger cAMP, which increases in concentration in response to stimulation by epinephrine (in muscle) or glucagon (in liver). Elevated [cAMP] initiates an enzyme cascade, in which a catalyst activates a catalyst, which activates a catalyst. Such cascades al low for large amplification of the initial signal (see pink boxes in Fig. 15–25). The rise in [cAMP] activates cAMP dependent protein kinase, also called protein kinase A (PKA). PKA then phosphorylates and activates phos phorylase b kinase, which catalyzes the phosphorylation of Ser residues in each of the two identical subunits of glycogen phosphorylase, activating it and thus stimulating glycogen breakdown. In muscle, this provides fuel for glycolysis to sustain muscle contraction for the fight-or-flight response signaled by epinephrine. In liver, glycogen breakdown counters the low blood glucose signaled by glucagon, releasing glucose. These different roles are reflected in subtle differences in the regulatory mechanisms in muscle and liver. The glycogen phosphorylases of liver and muscle are isozymes, en coded by different genes and differing in their regulatory properties.
In muscle, superimposed on the regulation of phos phorylase by covalent modification are two allosteric control mechanisms (Fig. 15–25). Ca2+, the signal for muscle contraction, binds to and activates phosphory lase b kinase, promoting conversion of phosphorylase b to the active a form. Ca2+ binds to phosphorylase b ki nase through its subunit, which is calmodulin (see Fig. 12–21). AMP, which accumulates in vigorously contracting muscle as a result of ATP breakdown, binds to and activates phosphorylase, speeding the release of glucose 1-phosphate from glycogen. When ATP levels are adequate, ATP blocks the allosteric site to which AMP binds, inactivating phosphorylase. When the muscle returns to rest, a second enzyme, phosphorylase a phosphatase, also called phospho protein phosphatase 1 (PP1), removes the phosphoryl groups from phosphorylase a, converting it to the less active form, phosphorylase b. Like the enzyme of muscle, the glycogen phosphorylase of liver is regulated hormonally (by phosphorylation/dephosphorylation) and allosterically. The dephosphorylated form is essentially inactive. When the blood glucose level is too low, glucagon (acting by the same cascade mechanism shown in Fig. 15–25) activates phosphorylase b kinase, which in turn converts phos phorylase b to its active a form, initiating the release of glucose into the blood. When blood glucose levels return to normal, glucose enters hepatocytes and binds to an inhibitory allosteric site on phosphorylase a. This binding also produces a conformational change that ex poses the phosphorylated Ser residues to PP1, which catalyzes their dephosphorylation and inactivates the phosphorylase (Fig. 15–26). The allosteric site for glu cose allows liver glycogen phosphorylase to act as its own glucose sensor and to respond appropriately to changes in blood glucose.
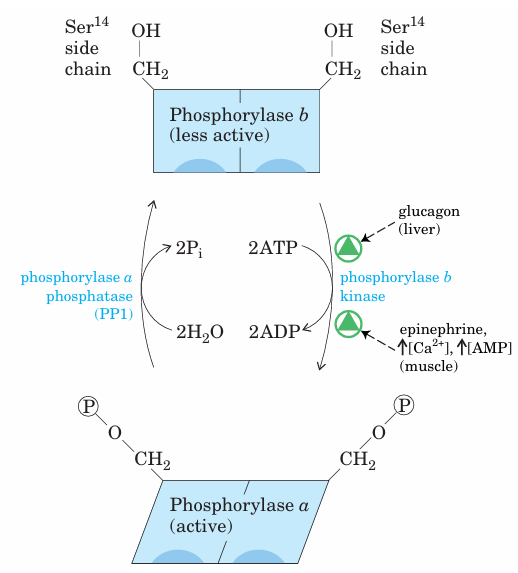
FIGURE 15–24 Regulation of muscle glycogen phosphorylase by covalent modification. In the more active form of the enzyme, phos phorylase a, Ser14 residues, one on each subunit, are phosphorylated. Phosphorylase a is converted to the less active form, phosphorylase b, by enzymatic loss of these phosphoryl groups, catalyzed by phos phorylase a phosphatase (PP1). Phosphorylase b can be reconverted (reactivated) to phosphorylase a by the action of phosphorylase b kinase.
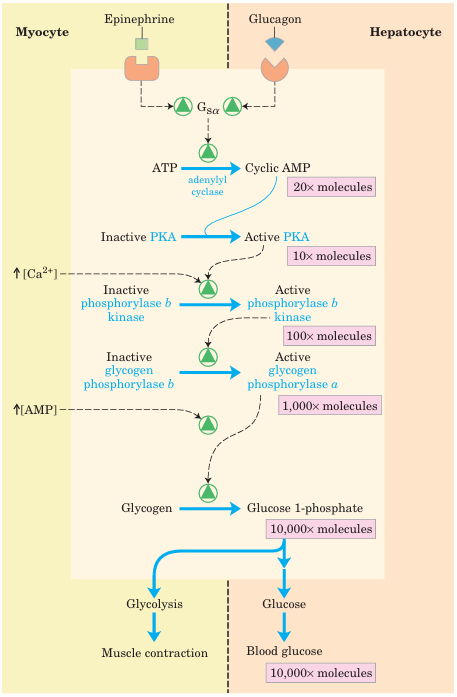
FIGURE 15–25 Cascade mechanism of epinephrine and glucagon action. By binding to specific surface receptors, either epinephrine acting on a myocyte (left) or glucagon acting on a hepatocyte (right) activates a GTP-binding protein Gsα (see Fig. 12–12). Active Gsα triggers a rise in [cAMP], activating PKA. This sets off a cascade of phosphorylations; PKA activates phosphorylase b kinase, which then activates glycogen phosphorylase. Such cascades effect a large amplification of the initial signal; the figures in pink boxes are probably low estimates of the actual increase in number of molecules at each stage of the cascade. The resulting breakdown of glycogen provides glucose, which in the myocyte can supply ATP (via glycolysis) for muscle contraction and in the hepatocyte is released into the blood to counter the low blood glucose.
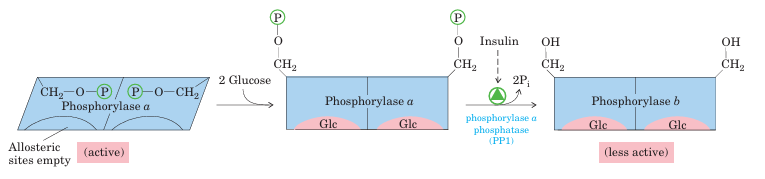
FIGURE 15–26 Glycogen phosphorylase of liver as a glucose sensor. Glucose binding to an allosteric site of the phosphorylase a isozyme of liver induces a conformational change that exposes its phosphorylated Ser residues to the action of phosphorylase a phosphatase 1(PP1).
This phosphatase converts phosphorylase a to phosphorylase b, sharply reducing the activity of phosphorylase and slowing glycogen breakdown in response to high blood glucose. Insulin also acts indirectly to stimulate PP1 and slow glycogen breakdown.
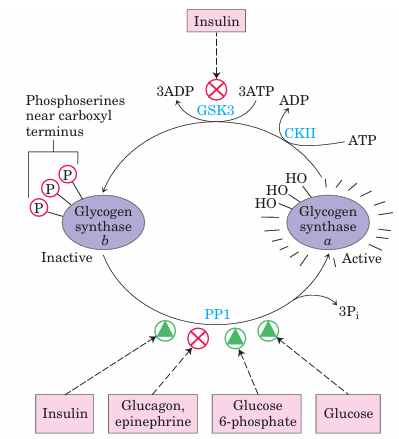
FIGURE 15–27 Effects of GSK3 on glycogen synthase activity. Glycogen synthase a, the active form, has three Ser residues near its carboxyl terminus, which are phosphorylated by glycogen synthase kinase 3 (GSK3). This converts glycogen synthase to the inactive (b) form (GSb). GSK3 action requires prior phosphorylation (priming) by casein kinase (CKII). Insulin triggers activation of glycogen synthase b by blocking the activity of GSK3 (see the pathway for this action in Fig. 12–8) and activating a phosphoprotein phosphatase (PP1 in muscle, another phosphatase in liver). In muscle, epinephrine activates PKA, which phosphorylates the glycogen-targeting protein GM (see Fig. 15–30) on a site that causes dissociation of PP1 from glycogen. Glucose 6-phosphate favors dephosphorylation of glycogen synthase by binding to it and promoting a conformation that is a good substrate for PP1. Glucose also promotes dephosphorylation; the binding of glucose to glycogen phosphorylase a forces a conformational change that favors dephosphorylation to glycogen phosphorylase b, thus relieving its inhibition of PP1.





















