Coordinated Regulation of Glycolysis and Gluconeogenesis:- Fructose 2,6-Bisphosphate Is a Potent Regulator of Glycolysis and Gluconeogenesis
The special role of liver in maintaining a constant blood glucose level requires additional regulatory mechanisms to coordinate glucose production and consumption. When the blood glucose level decreases, the hormone glucagon signals the liver to produce and release more glucose and to stop consuming it for its own needs. One source of glucose is glycogen stored in the liver; another source is gluconeogenesis. The hormonal regulation of glycolysis and gluconeogenesis is mediated by fructose 2,6-bisphosphate, an allosteric effector for the enzymes PFK-1 and FBPase-1 (Fig. 15–22):
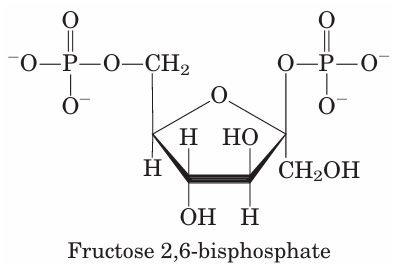
When fructose 2,6-bisphosphate binds to its allosteric site on PFK-1, it increases that enzyme’s affinity for its substrate, fructose 6-phosphate, and reduces its affinity for the allosteric inhibitors ATP and citrate. At the physiological concentrations of its substrates ATP and fructose 6-phosphate and of its other positive and negative effectors (ATP, AMP, citrate), PFK-1 is virtually inactive in the absence of fructose 2,6-bisphosphate. Fructose 2,6-bisphosphate activates PFK-1 and stimulates glycolysis in liver and, at the same time, inhibits FBPase-1, thereby slowing gluconeogenesis. Although structurally related to fructose 1,6 bisphosphate, fructose 2,6-bisphosphate is not an intermediate in gluconeogenesis or glycolysis; it is a regulator whose cellular level reflects the level of glucagon in the blood, which rises when blood glucose falls. The cellular concentration of fructose 2,6-bisphosphate is set by the relative rates of its formation and breakdown (Fig. 15–23a). It is formed by phosphorylation of fructose 6-phosphate, catalyzed by phosphofructoki nase-2 (PFK-2), and is broken down by fructose 2,6 bisphosphatase (FBPase-2). (Note that these en zymes are distinct from PFK-1 and FBPase-1, which catalyze the formation and breakdown, respectively, of fructose 1,6-bisphosphate.) PFK-2 and FBPase-2 are two distinct enzymatic activities of a single, bifunctional protein. The balance of these two activities in the liver, which determines the cellular level of fructose 2,6 bisphosphate, is regulated by glucagon and insulin (Fig. 15–23b). As we saw in Chapter 12 (p. 441), glucagon stimulates the adenylyl cyclase of liver to synthesize 3,5-cyclic AMP (cAMP) from ATP. Then cAMP activates cAMP-dependent protein kinase, which transfers a phosphoryl group from ATP to the bifunctional protein PFK-2/FBPase-2. Phosphorylation of this protein enhances its FBPase-2 activity and inhibits its PFK-2 activity. Glucagon thereby lowers the cellular level of fructose 2,6-bisphosphate, inhibiting glycolysis and stimulating gluconeogenesis. The resulting production of more glucose enables the liver to replenish blood glucose in response to glucagon. Insulin has the opposite effect, stimulating the activity of a phosphoprotein phosphatase that catalyzes removal of the phosphoryl group from the bifunctional protein PFK-2/FBPase-2, activating its PFK-2 activity, increasing the level of fructose 2,6-bisphosphate, stimulating glycolysis, and inhibiting gluconeogenesis.
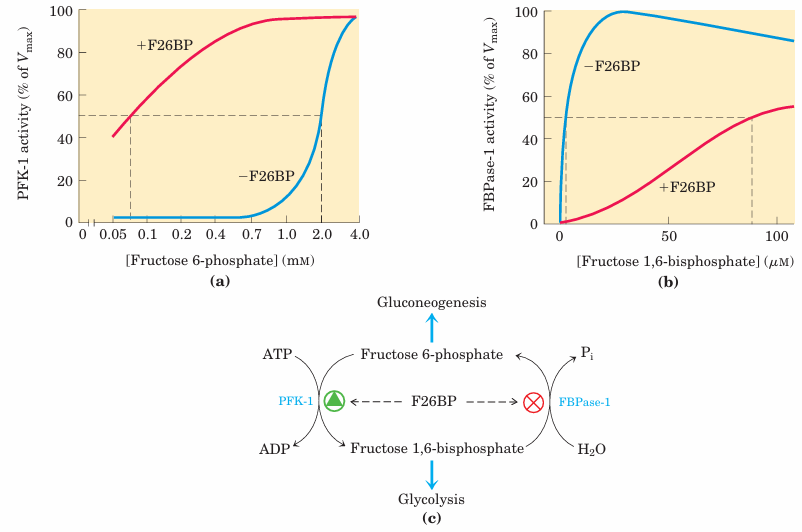
FIGURE 15–22 Role of fructose 2,6-bisphosphate in regulation of glycolysis and gluconeogenesis. Fructose 2,6-bisphosphate (F26BP) has opposite effects on the enzymatic activities of phosphofructoki nase-1 (PFK-1, a glycolytic enzyme) and fructose 1,6-bisphosphatase (FBPase-1, a gluconeogenic enzyme). (a) PFK-1 activity in the absence of F26BP (blue curve) is half-maximal when the concentration of fructose 6-phosphate is 2 mM (that is, K0.5 2 mM). When 0.13 µM F26BP is present (red curve), the K0.5 for fructose 6-phosphate is only 0.08 mM. Thus, F26BP activates PFK-1 by increasing its apparent affinity (Fig. 15–18) for fructose 6-phosphate. (b) FBPase-1 activity is inhibited by as little as 1 M F26BP and is strongly inhibited by 25 M. In the absence of this inhibitor (blue curve) the K0.5 for fructose 1,6 bisphosphate is 5 M, but in the presence of 25 M F26BP (red curve) the K0.5 is>70µM. Fructose 2,6-bisphosphate also makes FBPase-1 more sensitive to inhibition by another allosteric regulator, AMP. (c) Summary of regulation by F26BP.
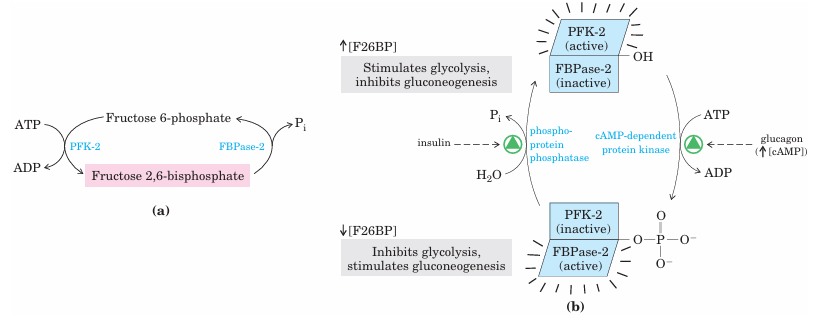
FIGURE 15–23 Regulation of fructose 2,6-bisphosphate level. (a)The cellular concentration of the regulator fructose 2,6-bisphosphate (F26BP) is determined by the rates of its synthesis by phosphofructo kinase-2 (PFK-2) and breakdown by fructose 2,6-bisphosphatase (FBPase-2). (b) Both enzymes are part of the same polypeptide chain, and both are regulated, in a reciprocal fashion, by insulin and glucagon. Here and elsewhere, arrows are used to indicate increasing (↑) and decreasing (↓) levels of metabolites.





















