






















تاريخ الفيزياء
علماء الفيزياء


الفيزياء الكلاسيكية
الميكانيك
الديناميكا الحرارية
الكهربائية والمغناطيسية
الكهربائية
المغناطيسية
الكهرومغناطيسية
علم البصريات
تاريخ علم البصريات
الضوء
مواضيع عامة في علم البصريات
الصوت
الفيزياء الحديثة
النظرية النسبية
النظرية النسبية الخاصة
النظرية النسبية العامة
مواضيع عامة في النظرية النسبية
ميكانيكا الكم
الفيزياء الذرية
الفيزياء الجزيئية
الفيزياء النووية
مواضيع عامة في الفيزياء النووية
النشاط الاشعاعي
فيزياء الحالة الصلبة
الموصلات
أشباه الموصلات
العوازل
مواضيع عامة في الفيزياء الصلبة
فيزياء الجوامد
الليزر
أنواع الليزر
بعض تطبيقات الليزر
مواضيع عامة في الليزر
علم الفلك
تاريخ وعلماء علم الفلك
الثقوب السوداء
المجموعة الشمسية
الشمس
كوكب عطارد
كوكب الزهرة
كوكب الأرض
كوكب المريخ
كوكب المشتري
كوكب زحل
كوكب أورانوس
كوكب نبتون
كوكب بلوتو
القمر
كواكب ومواضيع اخرى
مواضيع عامة في علم الفلك
النجوم
البلازما
الألكترونيات
خواص المادة
الطاقة البديلة
الطاقة الشمسية
مواضيع عامة في الطاقة البديلة
المد والجزر
فيزياء الجسيمات
الفيزياء والعلوم الأخرى
الفيزياء الكيميائية
الفيزياء الرياضية
الفيزياء الحيوية
الفيزياء العامة
مواضيع عامة في الفيزياء
تجارب فيزيائية
مصطلحات وتعاريف فيزيائية
وحدات القياس الفيزيائية
طرائف الفيزياء
مواضيع اخرى

The physiology of the eye
 المؤلف:
Richard Feynman, Robert Leighton and Matthew Sands
المؤلف:
Richard Feynman, Robert Leighton and Matthew Sands
 المصدر:
The Feynman Lectures on Physics
المصدر:
The Feynman Lectures on Physics
 الجزء والصفحة:
Volume I, Chapter 36
الجزء والصفحة:
Volume I, Chapter 36
 2024-04-06
2024-04-06
 2134
2134




+
-
20
We begin by talking not only about color vision, but about vision in general, just to remind ourselves about the interconnections in the retina. The retina is really like the surface of the brain. Although the actual picture through a microscope is a little more complicated looking than this somewhat schematized drawing, by careful analysis one can see all these interconnections. There is no question that one part of the surface of the retina is connected to other parts, and that the information that comes out on the long axons, which produce the optic nerve, are combinations of information from many cells. There are three layers of cells in the succession of function: there are retinal cells, which are the ones that the light affects, an intermediate cell which takes information from a single or a few retinal cells and gives it out again to several cells in a third layer of cells and carries it to the brain. There are all kinds of cross connections between cells in the layers.
We now turn to some aspects of the structure and performance of the eye. The focusing of the light is accomplished mainly by the cornea, by the fact that it has a curved surface which “bends” the light. This is why we cannot see clearly under water, because we then do not have enough difference between the index of the cornea, which is 1.37, and that of the water, which is 1.33. Behind the cornea is water, practically, with an index of 1.33, and behind that is a lens which has a very interesting structure: it is a series of layers, like an onion, except that it is all transparent, and it has an index of 1.40 in the middle and 1.38 at the outside. (It would be nice if we could make optical glass in which we could adjust the index throughout, for then we would not have to curve it as much as we do when we have a uniform index.) Furthermore, the shape of the cornea is not that of a sphere. A spherical lens has a certain amount of spherical aberration. The cornea is “flatter” at the outside than is a sphere, in just such a manner that the spherical aberration is less for the cornea than it would be if we put a spherical lens in there! The light is focused by the cornea-lens system onto the retina. As we look at things that are closer and farther away, the lens tightens and loosens and changes the focus to adjust for the different distances. To adjust for the total amount of light there is the iris, which is what we call the color of the eye, brown or blue, depending on who it is; as the amount of light increases and decreases, the iris moves in and out.
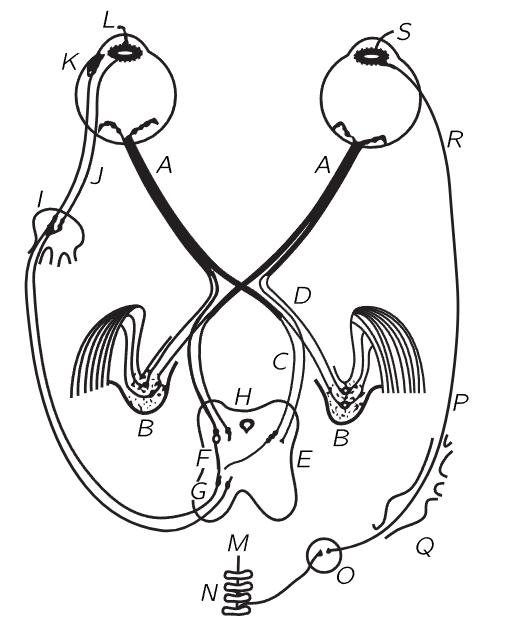
Fig. 36–3. The neural interconnections for the mechanical operation of the eyes.
Let us now look at the neural machinery for controlling the accommodation of the lens, the motion of the eye, the muscles which turn the eye in the socket, and the iris, shown schematically in Fig. 36–3. Of all the information that comes out of the optic nerve A, the great majority is divided into one of two bundles (which we will talk about later) and thence to the brain. But there are a few fibers, of interest to us now, which do not run directly to the visual cortex of the brain where we “see” the images, but instead go into the mid-brain H. These are the fibers which measure the average light and make adjustment for the iris; or, if the image looks foggy, they try to correct the lens; or, if there is a double image, they try to adjust the eye for binocular vision. At any rate, they go through the mid-brain and feed back into the eye. At K are the muscles which run the accommodation of the lens, and at L another one that runs into the iris. The iris has two muscle systems. One is a circular muscle L which, when it is excited, pulls in and closes down the iris; it acts very rapidly and the nerves are directly connected from the brain through short axons into the iris. The opposite muscles are radial muscles, so that, when the things get dark and the circular muscle relaxes, these radial muscles pull out. Here we have, as in many places in the body, a pair of muscles which work in opposite directions, and in almost every such case the nerve systems which control the two are very delicately adjusted, so that when signals are sent in to tighten one, signals are automatically sent in to loosen the other. The iris is a peculiar exception: the nerves which make the iris contract are the ones we have already described, but the nerves which make the iris expand come out from no one knows exactly where, go down into the spinal cord back of the chest, into the thoracic sections, out of the spinal cord, up through the neck ganglia, and all the way around and back up into the head in order to run the other end of the iris. In fact, the signal goes through a completely different nervous system, not the central nervous system at all, but the sympathetic nervous system, so it is a very strange way of making things go.
We have already emphasized another strange thing about the eye, that the light-sensitive cells are on the wrong side, so that the light has to go through several layers of other cells before it gets to the receptors—it is built inside out! So, some of the features are wonderful and some are apparently stupid.
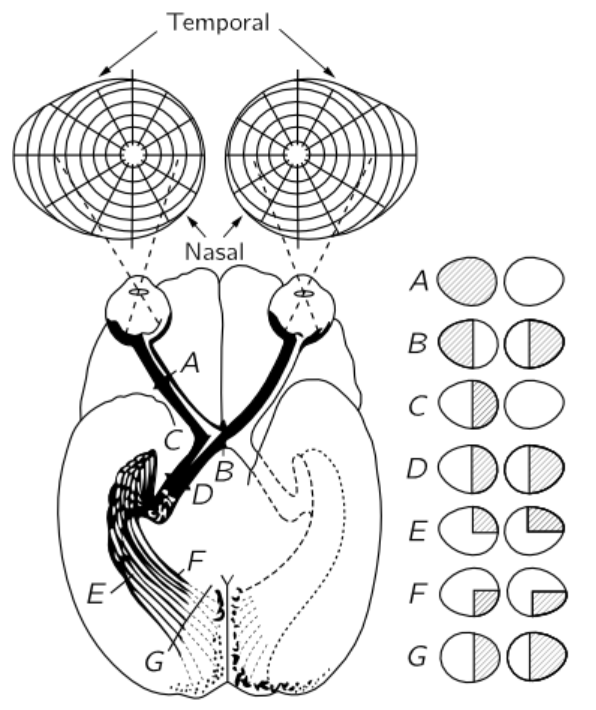
Fig. 36–4. The neural connections from the eyes to the visual cortex.
Figure 36–4 shows the connections of the eye to the part of the brain which is most directly concerned with the visual process. The optic nerve fibers run into a certain area just beyond D, called the lateral geniculate, whereupon they run out to a section of the brain called the visual cortex. Notice that some of the fibers from each eye are sent over to the other side of the brain, so the picture formed is incomplete. The optic nerves from the left side of the right eye run across the optic chiasma B, while the ones on the left side of the left eye come around and go this same way. So, the left side of the brain receives all the information which comes from the left side of the eyeball of each eye, i.e., on the right side of the visual field, while the right side of the brain sees the left side of the visual field. This is the manner in which the information from each of the two eyes is put together in order to tell how far away things are. This is the system of binocular vision.
The connections between the retina and the visual cortex are interesting. If a spot in the retina is excised or destroyed in any way, then the whole fiber will die, and we can thereby find out where it is connected. It turns out that, essentially, the connections are one to one—for each spot in the retina there is one spot in the visual cortex—and spots that are very close together in the retina are very close together in the visual cortex. So, the visual cortex still represents the spatial arrangement of the rods and cones, but of course much distorted. Things which are in the center of the field, which occupy a very small part of the retina, are expanded over many, many cells in the visual cortex. It is clear that it is useful to have things which are originally close together, still close together. The most remarkable aspect of the matter, however, is the following. The place where one would think it would be most important to have things close together would be right in the middle of the visual field. Believe it or not, the up-and-down line in our visual field as we look at something is of such a nature that the information from all the points on the right side of that line is going into the left side of the brain, and information from the points on the left side is going into the right side of the brain, and the way this area is made, there is a cut right down through the middle, so that the things that are very close together right in the middle are very far apart in the brain! Somehow, the information has to go from one side of the brain to the other through some other channels, which is quite surprising.
The question of how this network ever gets “wired” together is very interesting. The problem of how much is already wired and how much is learned is an old one. It used to be thought long ago that perhaps it does not have to be wired carefully at all, it is only just roughly interconnected, and then, by experience, the young child learns that when a thing is “up there” it produces some sensation in the brain. (Doctors always tell us what the young child “feels,” but how do they know what a child feels at the age of one?) The child, at the age of one, supposedly sees that an object is “up there,” gets a certain sensation, and learns to reach “there,” because when he reaches “here,” it does not work. That approach probably is not correct, because we already see that in many cases there are these special detailed interconnections. More illuminating are some most remarkable experiments done with a salamander. (Incidentally, with the salamander there is a direct crossover connection, without the optic chiasma, because the eyes are on each side of the head and have no common area. Salamanders do not have binocular vision.) The experiment is this. We can cut the optic nerve in a salamander and the nerve will grow out from the eyes again. Thousands and thousands of cell fibers will thus re-establish themselves. Now, in the optic nerve the fibers do not stay adjacent to each other—it is like a great, sloppily made telephone cable, all the fibers twisting and turning, but when it gets to the brain, they are all sorted out again. When we cut the optic nerve of the salamander, the interesting question is, will it ever get straightened out? The answer is remarkable: yes. If we cut the optic nerve of the salamander and it grows back, the salamander has good visual acuity again. However, if we cut the optic nerve and turn the eye upside down and let it grow back again, it has good visual acuity all right, but it has a terrible error: when the salamander sees a fly “up here,” it jumps at it “down there,” and it never learns. Therefore, there is some mysterious way by which the thousands and thousands of fibers find their right places in the brain.
This problem of how much is wired in, and how much is not, is an important problem in the theory of the development of creatures. The answer is not known, but is being studied intensively.
The same experiment in the case of a goldfish shows that there is a terrible knot, like a great scar or complication, in the optic nerve where we cut it, but in spite of all this the fibers grow back to their right places in the brain.
In order to do this, as they grow into the old channels of the optic nerve they must make several decisions about the direction in which they should grow. How do they do this? There seem to be chemical clues that different fibers respond to differently. Think of the enormous number of growing fibers, each of which is an individual differing in some way from its neighbors; in responding to whatever the chemical clues are, it responds in a unique enough way to find its proper place for ultimate connection in the brain! This is an interesting—a fantastic—thing. It is one of the great recently discovered phenomena of biology and is undoubtedly connected to many older unsolved problems of growth, organization, and development of organisms, and particularly of embryos.
One other interesting phenomenon has to do with the motion of the eye. The eyes must be moved in order to make the two images coincide in different circumstances. These motions are of different kinds: one is to follow something, which requires that both eyes must go in the same direction, right or left, and the other is to point them toward the same place at various distances away, which requires that they must move oppositely. The nerves going into the muscles of the eye are already wired up for just such purposes. There is one set of nerves which will pull the muscles on the inside of one eye and the outside of the other, and relax the opposite muscles, so that the two eyes move together. There is another center where an excitation will cause the eyes to move in toward each other from parallel. Either eye can be turned out to the corner if the other eye moves toward the nose, but it is impossible consciously or unconsciously to turn both eyes out at the same time, not because there are no muscles, but because there is no way to send a signal to turn both eyes out, unless we have had an accident or there is something the matter, for instance if a nerve has been cut. Although the muscles of one eye can certainly steer that eye about, not even a Yogi is able to move both eyes out freely under voluntary control, because there does not seem to be any way to do it. We are already wired to a certain extent. This is an important point, because most of the earlier books on anatomy and psychology, and so on, do not appreciate or do not emphasize the fact that we are so completely wired already—they say that everything is just learned.
 الاكثر قراءة في الفيزياء الحيوية
الاكثر قراءة في الفيزياء الحيوية
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















