آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Other Types of Roots and Root Modifications |


|
|
Read More
Date: 14-11-2016
Date: 21-11-2016
Date: 14-11-2016
|
Other Types of Roots and Root Modifications
The type of root just described is the most common, generalized type, comprising some or all of the roots of most plants. In other species, some roots are modified and carry out different roles in the plant's survival. Various structures and organizations are selectively advantageous for various functions.
STORAGE ROOTS
Rods frequently provide long-term storage for carbohydrates that accumulate during summer photosynthesis. This occurs only in perennial plants, because the carbohydrates are used in the production of new shoots or flowers in the spring, when photosynthesis is impossible owing to the lack of leaves. Annual plants can survive without such storage capacity.
Deciduous perennials also store significant amounts of nutrients in the stem during winter, but the roots offer certain advantages. Being subterranean, they are less available as food than are swollen, highly nutritious, easily visible stems. Roots also have a much more stable environment, subjected to less extreme changes in temperature and humidity; this may be important for the survival of the storage parenchyma cells. In many perennials, m& of the stem is annual, dying back to a few nodes located below ground at the top of the root It seems less economical in these species to winterproof the shoot than to replace it using nutrients stored in the roots.
PROP ROOTS
The stem of a monocot can become wider, with more vascular bundles, if it can produce adventitious roots that extend to the soil. In some cases, the roots are capable of extensive growth through the air: In many palms, the exposed roots can be 20 to 50 cm long (Fig. 1a). In the screwpine (Pandanus, a monocot) they are often 3 or 4 meters long, and the roots may grow through the air for months before reaching the ground (Fig. 1b). dramatic examples are found at the bases of corn and many other grasses.




FIGURE 1: (a) Many monocots like this palm (Ptychosperma) produce adventitious roots near the base of the stem These provide both extra absorptive capacity and extra stability. (b and c) Screwpine is able to produce extremely long adventitious pep roots that not only stabilize the large, heavy tank but also bring water and minerals into the stem (d) In older plants of screwpine, even the branches produce adventitious roots. This water and mineral supply to the branches bypasses the trunk, where the vascular bundles are already conducing at full capacity.
Once the prop roots do make contact with the soil, they transport additional nutrients and water to the stem. Just as importantly, they contract slightly and place some tension on the stem, thus acting as stabilizers, much like guy wires on telephone poles. If the roots undergo secondary growth and become woody, they can be extremely strong supports, permitting a branch to extend even farther from the trunk without breaking or sagging (Fig. 2). In banyan trees (dicots of the genus Ficus), the prop roots and branches can spread and produce massive trees many meters in diameter.

FIGURE 2: Banyan trees produce adventitious roots which, as in the monocots, provide increased support and absorptive capacity. Because the giant branches are supported along their entire length, they can become much larger and more extensive than branches that are supported only at the point of attachment to the trunk.
Mangroves also have prop roots, but these seem to be selectively advantageous for other reasons. The plants grow in intertidal marshes and are subjected to powerful water currents during storms and even normal tide changes; the bracelike prop roots provide much more stability than a taproot system would. In addition, the aerial portion of the prop root is covered with numerous air chambers-lenticels-and its cortex is a wide aerenchyma similar to that in the knees of bald cypress. The subterranean portion of the root grows in a stagnant muck that has little or no oxygen; it is able to respire only because the aerenchyma permits the rapid diffusion of oxygen from the aerial lenticels to the sub-merged root tissues. If the roots were entirely subterranean, respiration would be extremely reduced.
AERIAL ROOTS OF ORCHIDS
Many orchids are epiphytic, living attached to the branches of trees. Their roots spread along the surface of the bark and often dangle freely in the air. Although these plants live in rainforests, the orchids are actually adapted to drought conditions. In the few hours when rain does not fall, the air and bark become dry and could easily pull water out of the orchid's roots if there were no water-conserving mechanism. The root epidermis, called a velamen in these orchids, is composed of several layers of large dead cells that are white in appearance (Fig. 3). Apparently the velamen acts as a water-proof barrier, not permitting water to leave the sides of the root.

FIGURE 3: Aerial roots of orchids, which grow along the surface of tree bark, are covered by a white velamen that helps retain the water absorbed by the green root tip.
CONTRACTILE ROOTS
In Oxalis, Gladiolus, Crinum, and other plants (many with bulbs), roots undergo even more contraction than prop roots do. After extending through the soil and becoming firmly anchored, the uppermost portions begin to contract (Fig. 4). Because the root is firmly fixed to the soil, the stem is pulled downward so that the base of the shoot is either kept at soil level or, in the case of bulbs, actually buried deeper. The contraction is caused by changes in the shape of cortical cells: They simultaneously expand radially and shorten, losing as much as one half to two thirds of their height. The vascular tissues buckle and become undulate but are able to continue conducting.
Contractile roots may be more common than is generally appreciated. Many seeds germinate at or near the soil surface; root contraction may be the means by which the shoot becomes anchored in the soil. In bulbs, corms, rhizomes, and other subterranean stems, contractile roots can be important in keeping the stems at the proper depth.

FIGURE 4: The roots of this hyacinth have contracted, causing their surfaces to become wrinkled.
MYCORRHIZAE
The roots of most species of seed plants (at least 80%) have a symbiotic relationship with soil fungi in which both organisms benefit. The associations are known as mycorrhizae (sing.: mycorrhiza), and two main types of relationships are known. In nearly all woody forest plants, an ectomycorrhizal relationship exists, in which the fungal hyphae (slender threadlike cells) penetrate between the outermost root cortex cells but never invade the cells themselves. Herbaceous plants have an endomycorrhizal association, in which the hyphae penetrate the root cortex as far as the endodermis; they are able to pass through the walls of the cortical cells but cannot pass the Casparian strip (Fig. 5). They invade the cell but do not break the host plasma membrane or vacuole membrane. Inside the cell they branch repeatedly, forming a small structure called an arbuscule; this fills with granules of phosphorus which later disappear as the phosphorus is absorbed by the plant.

FIGURE 5: (a) Diagram of endomycorrhizae showing hyphae penetrating the root cortex and forming arbuscules and vesicles. (b) Root cells filled with fungal hyphae in the form of arbuscules. You can tell that this is a symbiotic fungus rather than a pathogenic one because the plant cells are healthy, with large normal nuclei (X 220). (c) Roots of beech encased in ectomycorrhizae. The' presence of the fungus stimulates the roots to be short and broad, (d) Transverse section of an ectomycorrhizal root showing that the fungal filaments are restricted to just the outermost part of the root; they do not penetrate the cortex.
Other hyphae fill with membranous vesicles. The plant cells lack starch grains, presumably because the sugars are being transferred to the fungus. The fungus is unable to live without the sugars from the plant, and, in many cases, the plant is severely stunted if the fungus is killed. Apparently, this is a critical means of absorbing phosphorus into the roots, the mycorrhizal fungi being much more effective than root hairs (Fig. 6).

FIGURE 6: The ability of barley roots to absorb phosphorus from solutions of different concentrations is shown on this graph. Both sterilized roots (fungi killed) and natural roots (fungi still alive) absorb phosphorus more rapidly from the solutions with the high concentrations of phosphorus, but at every concentration the roots with mycorrhizal fungi absorb much more phosphorus than the sterile roots.
All the modified roots mentioned to this point—storage roots, prop roots, contractile roots, aerial roots of orchids, and mycorrhizae-associated roots-have a structure virtually identical to that of the more common anchoring/absorbing roots described above. All grow by an apical meristem, and a cross-section through a mature region of each reveals epidermis, cortex, endodermis, pericycle, and vascular tissue. In nitrogen-fixing nodules and haustoria, however, the structure is highly modified.
ROOT NODULES AND NITROGEN FIXATION
For most plants, the scarcity of nitrogenous compounds in the soil is one of the main growth-limiting factors. Although nitrogen is abundant in the air (78% of the atmosphere is N2), plants have no enzyme systems that can use that nitrogen. Only some prokaryotes can use N2 by incorporating it into their bodies as amino acids and nucleotides; when they die and decompose, the nitrogenous compounds are available to plants. The chemical process of converting atmospheric nitrogen into usable compounds is nitrogen fixation.
In a small number of plants, especially legumes, a symbiotic relationship has evolved with nitrogen-fixing bacteria of the genus Rhizobium. Bacteria free in the soil secrete a substance that causes root hairs to curl sharply; the bacterium then attaches to the convex side of the hair and pushes into the cell by means of a tube-like invagination of the plant cell i wall. The tube is an infection thread, and the bacterium sits in it. The infection thread extends all the way into the root's inner cortex, where adjacent cortical cells undergo j mitosis and form a root nodule (Fig. 7); the bacterium is released from the infection thread and enters the host cell cytoplasm, where it proliferates rapidly, filling the host cells with bacterial cells (known as bacteroids) capable to converting N2 into nitrogenous compounds that are released to the host cells. Energy for this process is supplied by sugars from tie legume root cells, so both the Rhizobium and the legume benefit.
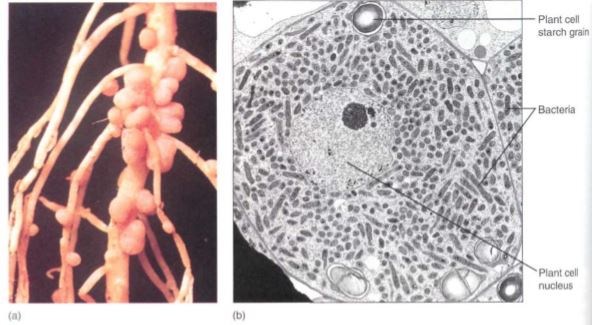
FIGURE 7: (a) The nodules on these alfalfa roots contain bacteria capable of absorbing atmospheric nitrogen and converting it to ammonia, which the plant can use to make amino acids. (Hugh Spencer/Photo Researchers, Inc.) (b) Bacteria in an infected root nodule cell of cowpea, Vigna unguiculata (X 5900). (Courtesy of E. H. Newcomb, University ofWisconsin, Madison)
The nodule may remain rather simple or may become complex, with a meristematic region, vascular tissue, and endodermis. Such nodules are able to function for extended lengths of time. Numerous metabolic changes have also evolved: The critical bacterial enzymes are sensitive to oxygen, being immediately poisoned by even traces of free oxygen. But it is the plant, not the bacterium, that produces a special chemical-leghemoglobin- that binds to oxygen and protects the bacterial enzymes from oxygen. These root nodules represent a sophisticated symbiosis; without the bacterium the complex development of nodules does not occur.
This process is often described thus: "The bacterium supplies the plant with nitrogenous compounds and in return the plant gives it sugars." This inaccurately suggests voluntay action, choice, and decision-making on the part of both organisms. Instead, think in terms of natural selection: Because the bacterium receives sugars and other nutrients from the plant, any genetic mutation in the bacterium that makes the plant healthier and more vigorous is beneficial to the bacterium as well, whereas any mutation that is harmful to the plant-such as a bacterial plasma membrane that does not allow nitrogenous compounds .
to move to the plant—is deleterious to the bacterium. The same principle applies to the plant: Genetic mutations that aid the bacterium ultimately benefit the plant as well. We hypothesize that these organisms will continue to co-evolve and become more fully co-adapted, but this hypothesis will have to be checked by someone a few million years from now.
Working backward, we can imagine less sophisticated legumes and Rhizobium bacteria that did not work so well together but evolved to the present state as mutations occurred and survived by natural selection. Certain critical mutations must be rare, or perhaps several unusual mutations must occur almost simultaneously in plant and bacterium; this hypothesis is based on the observation that, although this symbiosis would probably be beneficial to virtually all plant species, only a few have it.
HAUSTORIAL ROOTS OF PARASITIC FLOWERING PLANTS
A number of angiosperms are parasites on other plants; because their substrate is the body of another plant, a normal root system would not penetrate the host or absorb materials from it effectively. Consequently, the roots of parasitic plants have become highly modified and are known as haustoria (Fig. 8). In most cases, very little rootlike structure remains. Parasitism has evolved several times, so the structures that are termed haustoria ae not all related to each other and generalizations are difficult. However, haustoria typically must adhere firmly to their host either by secreting an adhesive or by growing around a small branch or root. Penetration occurs either by forcing a shaft of cells through the host dermal system or by expanding the haustorium radially, cracking the host epidermis. Air penetration, cells of the parasite make contact with the host xylem. In many cases both host and parasite cells divide and proliferate into an irregular mass of parenchyma, and then a column of cells differentiates into a series of vessel elements. This results in a continuous vessel from host to parasite constructed of cells of both. Surprisingly, many parasites attack only the xylem; they do not draw sugars from the host but carry out their own photosynthesis. Others do contact both xylem and phloem and perform little or no photosynthesis.
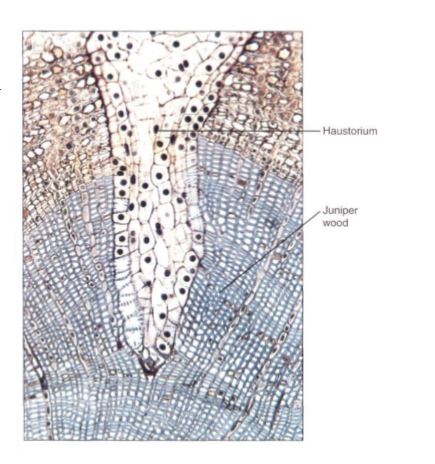
FIGURE 8: Transverse section of a branch of juniper being attacked by the haustorium (modified root) of a mistletoe (Phoradendron). The wood of the host juniper has been stained blue, the bark is tan. The haustorium is able to draw water and nutrients from the host vascular tissue.
For such parasites, mutations that result in the loss of leaves or vegetative stems are not disadvantageous. In the case of Tristerix, the parasite spends much of its life as nothing more than a diffuse "root" system embedded in the host .
The typical organization of a root would be nonfunctional in haustoria: The root cap, root apical meristem, and cortex would all prevent the vascular tissues of the parasite from contacting the appropriate tissues of the host. Conversely, haustoria are completely inadequate for growth in soil, however rich and moist it might be. For each substrate and each microhabitat, specific adaptations are selectively advantageous.



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
نحو شراكة وطنية متكاملة.. الأمين العام للعتبة الحسينية يبحث مع وكيل وزارة الخارجية آفاق التعاون المؤسسي
|
|
|