























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Growth Phase of The Cell Cycle
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 18-10-2016
18-10-2016
 2926
2926




+
-
20
Growth Phase of The Cell Cycle
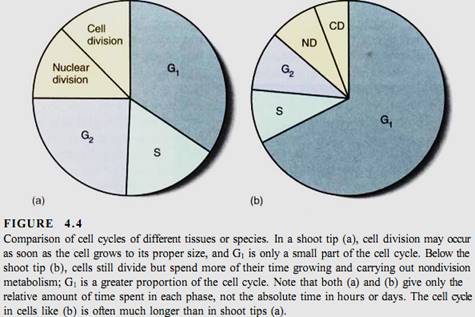
In the 1800s, when the cell cycle was first being studied intensively, researchers gave greatest attention to the division activities because many events could be identified. They assumed that between divisions cells were "resting," so the growth phase was called the resting phase, or interphase (Fig. 4.4). We now know that the cell is actually most active during interphase, and it is possible to detect three distinct phases within interphase: G1, S, and G2.
G1 PHASE
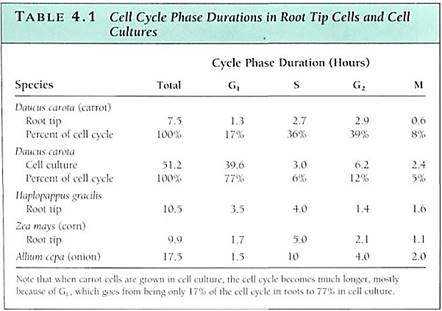
such as some algae, the cell cycle may be as brief as several hours; short cycle times also occur in rapidly growing embryos and roots. On the other hand, cell cycle times of 2 to 3 days or even weeks or months are not unusual in tissues or plants that grow slowly. During winter dormancy, the cell cycle may last from autumn until spring; when dormancy is broken and metabolism accelerates, the cell cycle may last only a few hours. The actual length of time that any cell spends in G1 is similarly variable. Although there are exceptions, in general G1 is the longest part of the cell cycle (Table 4.1). Once a cell undergoes cell cycle arrest, stops dividing, and begins to differentiate and mature, it may enter a state similar to G1 and remain in it for the rest of its life.
S PHASE
During the S (or synthesis) phase, the genes in the nucleus are replicated . A gene is a polymer of nucleotides, and each gene has a unique sequence of nucleotides. Although many genes may have similar sizes, different genes never have exactly the same sequence of nucleotides. It has been estimated that many higher plants and animals need about 100,000 types of genes to store all the information required to make the proper enzymes, structural proteins, and hormones necessary for the organism's life. The whole complex of genes for an organism is its genome.
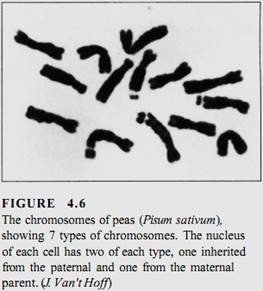
Obviously, if each individual gene floated around the cell or nucleus as an independent piece of DNA, then during division, it would be extraordinarily difficult to find each of those 200,000 genes (100,000 originals and 100,000 copies) and make certain that each daughter cell received one of each (Fig. 4.5). This is not a problem, however, because all genes are attached to other genes by short pieces of linking DNA. Thousands of genes can be attached together in a linear sequence; the whole structure is a chromosome (Fig. 4.6). Although it might seem reasonable for all genes to link together in a single chromosome, such a long chromosome would probably become hopelessly tangled. Only a low plants have as few as two chromosomes for their genome; most have between 5 and 30 chromosomes (Table 4.2).
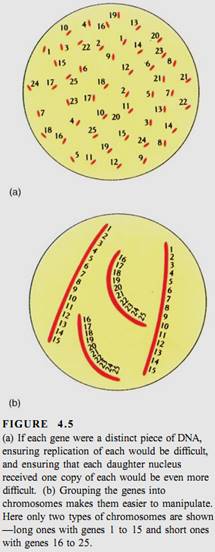
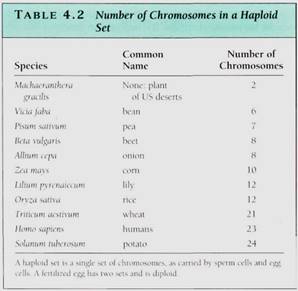
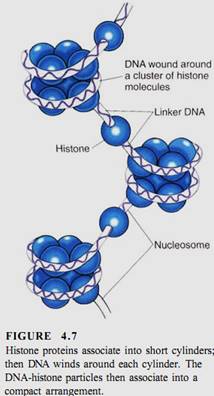
Even with 20 different chromosomes, an organism with 100,000 genes would have an average of 5000 genes on each chromosome, resulting in long pieces of DNA that might break if unprotected. In onion, the DNA in each nucleus is 10.5 meters long when all DNA molecules are measured; in lilies it is 21.8 meters long. In eukaryotes a special class of proteins called histones complexes with the DNA and gives it both protection and structure (Fig. 4.7). Chromosomes also have another structural feature, the centromere, which is usually located near the center of the chromosome (Fig. 4.8).

During the S phase, linking pieces of DNA as well as genes are replicated and new histone molecules complex with the new DNA. Thus entire chromosomes, not just DNA, are replicated (Fig. 4.9). Once replication is finished, the duplicate DNA molecules remain attached at the centromere. Although it would be justifiable to call this a "double chromo-some" or a "pair of chromosomes," it is also called a chromosome. Now that we know how a chromosome changes during S phase, we call each half of the doubled chromosome a chromatid. After S phase each chromosome has two identical chromatids; before S phase each chromosome has just one chromatid. It is important to remember that a chromosome after S phase is twice as large as it was before S phase.
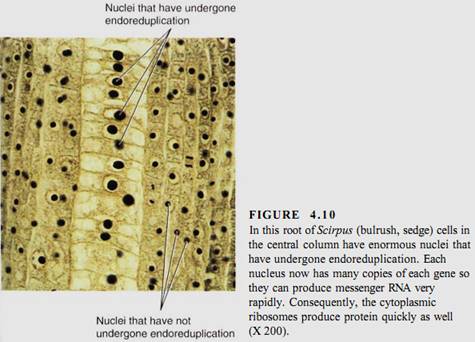
Although many cells stop in Gi when they cease dividing and begin to mature, most plant cells enter S phase and replicate their DNA before they begin to differentiate. This may involve just a single cycle of DNA replication, resulting in a nucleus that is twice as large as would be expected, or it may continue for many rounds of DNA synthesis and the nucleus becomes gigantic (Fig. 4.10). This process is endoreduplication, estimated to occur in 80% of all maturing plant cells (Table 4.3). The resulting nucleus has many copies of each gene-as many as 8192 copies in some cells of kidney beans (Phaseolus vulgaris). Endoreduplication occurs most often in hairs, glandular cells, and other cells that must have an extremely rapid, intense metabolism. The normal complement of two copies of each gene does not seem to make messenger RNA rapidly enough for such active cells. Such cells are also rich in ribosomes and produce large amounts of protein.
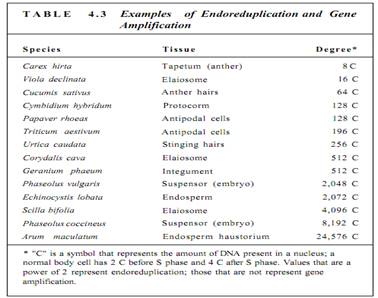

Gene amplification is similar to endoreduplication but involves only some genes that are repeatedly replicated. The amplified genes are those needed for the specialized metabolism of the mature cell. For example, as a protein-rich seed develops, its cells need large quantities of the messenger RNA that codes (or the storage protein, and those genes are amplified. But it would be a waste of energy and resources and deleterious to the plant to replicate all the other genes as well. The most extreme case of gene amplification known is in Arum maculatum (a relative of Philodendron): Each nucleus has enough DNA lor 24,576 copies of every gene. We do not know how many genes are being amplified and how many are present as only two copies per nucleus, so we cannot compute the actual number of copies of each amplified gene, but it is certainly more than 24,576.
G PHASE
synthesized, and the cell is believed to produce proteins necessary for processing chromosomes and breaking down the nuclear envelope. In cultured animal tissue, if a cell whose nucleus is dividing is forced to fuse with one in G1, the second cell's nucleus also begins the first steps of nuclear division. This is evidence that during G2, the first cell produced factors necessary to start nuclear division and that these factors are located in the cytoplasm. G1, S, and G2 constitute the interphase portion of the cell cycle. After G2, division can occur.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















