آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Membrane Transport |


|
|
Read More
Date: 15-10-2015
Date: 28-10-2015
Date: 23-10-2015
|
Membrane Transport
Biological membranes are the structures that divide biological space into functional aqueous compartments: cells and their organelles. To allocate hundreds of different solutes to their proper locations, cells equip their various membranes with multiple transport mechanisms, some simple and some complex. (Use of membrane vesicles for endocytosis and exocytosis is covered elsewhere.)
Membrane Diffusion
The simplest kind of transport is the unassisted diffusion of solutes across membranes (see Figure 1a). The kinds of molecules that transit in this fashion are more soluble in oil than water and so readily dissolve in and then spontaneously traverse the nonpolar lipid core of the membrane bilayer. Among these diffusible lipid-soluble molecules are steroid hormones, many kinds of drugs, the oxygen that cells respire, and the carbon dioxide they expire.
Motion of all kinds must be impelled by some form of energy. In the case of simple membrane diffusion, movement across the bilayer is a random walk driven by the kinetic (heat) energy provided by the collisions of the solute with surrounding molecules. This is the Brownian motion that agitates all molecules. Random diffusion causes the solute molecules to end up at equal concentrations on the two sides of the membrane no matter how great the initial difference (gradient) was. Solute transport by these means is thus said to be downhill.
Membrane Channels
Most cellular solutes have a polar chemical structure and are therefore strongly attracted to water. Consequently, these water-soluble molecules tend not to enter the lipid core of the membrane readily; indeed, the bilayer is designed keep them from doing so. To transport these solutes across the barrier, membranes are equipped with a variety of special protein structures.
The simplest way to convey water-soluble solutes across membranes is through channels: membrane-spanning proteins with central pores. By these means, selected solutes diffuse downhill across membranes, passing single file along a narrow column of water molecules in these pores. Driven by random Brownian diffusion, a solute will ultimately reach an equal concentration in the two aqueous compartments.
Channels can discriminate among solutes. They use the diameter of their pores as a sieve and place critical amino acid side chains along the pore to give it the proper shape and chemical profile. As a result, different channels strongly prefer Na+ or K+ or Ca++ or H+ and pass cations much better than anions; another channel is specialized for Cl-. Some channel families conduct larger solutes into cells; for example, nutrients like amino acids or sugars There are also elaborate channels that enable newly synthesized polypeptide chains to slither across the membrane in an extended conformation; they then fold up into mature proteins on the other side. This is the basis for protein secretion.
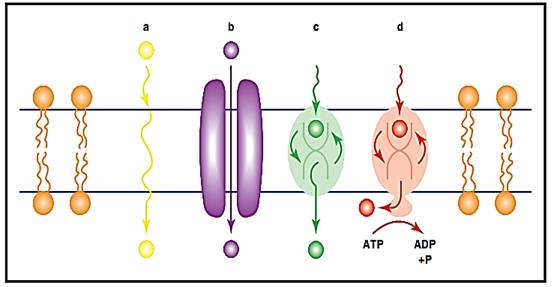
Figure 1. Modes of membrane transport. a) Some solute molecules can diffuse unassisted through the lipid bilayer. b) Certain solute molecules can diffuse though the aqueous pore of a specific channel protein. c) Reciprocating transporters can convey selected solute molecules across the bilayer by means of a fluctuating change in their shape. d) The energy released by the breakdown of ATP molecules can be coupled by a transport protein to pumping specific solute molecules against their concentration gradient.
Among the most interesting kinds of channels are those along the axon or at the synapses of nerve cells. When turned on for a few milliseconds by a stimulus, these proteins allow small numbers of Na+ or K+ to diffuse across the membrane. These pores open as the result of a change in the structure of the channel protein, typically brought about by one of two mechanisms: (1) the association of a specific neurotransmitter molecule to a binding site on the channel protein (at the synapse); or (2) a change in the electrical field across the membrane (along the axon). In both cases, the consequent movement of ions through these activated channels alters the electrical charge across the membrane, causing channels nearby to open. This electrical cascade is what propagates excitation along a nerve.
Reciprocating Transporters
Animal cells have membrane transporters that “carry” specific nutrients down their concentration gradients. These transport proteins work like an engine with a four-step cycle. The first step is for the membrane-bound transporter to bind a solute molecule outside of the cell. Each kind of transporter is selective for a specific solute family, just as an enzyme acts only upon certain substrates. Discrimination comes from the close-fitting association of the solute with a pocket in the protein, like a key in a lock or a hand in a glove. The shape of the transporter molecule fluctuates constantly; driven, once again, by Brownian (thermal) motion within the fluid membrane. In the second step, the right twists and turns cause the protein to “swallow” the solute, reorienting it from the outside to the interior of the cell. Third, the solute diffuses away from the transporter into the cytoplasm. Finally, the empty transporter reorients to its initial shape, so that its solute binding site again addresses the cell exterior. The protein is then ready for the next cycle.
Such transporters convey metabolites such as glucose and amino acids in the bloodstream (where they are at high concentration) to the cell’s cytoplasm (where they are being consumed). Their activity can be regulated according to need, often through the action of hormones. For example, when excess glucose is available, the hormone insulin is released from the pancreas into the bloodstream. This hormone signals muscle cells to bring their glucose transporters into play; these convey the sugar into the cells, returning the blood glucose level to normal.
This mode of transport utilizes the energy of Brownian motion in two ways: molecular collisions not only propel solute molecules up to and away from the transporter but also drive the protein to change its shape, back and forth, between its two functional orientations. No matter how well designed, these reciprocating transporters can only equalize the concentration of a solute between the two compartments the membrane separates. That is, random motion cannot gather up every molecule of a nutrient outside a cell or expel every molecule of waste from the interior of a cell. If the cell needs to take up or pump out a solute beyond its equal distribution, it must apply energy. The methods of transport just described are therefore referred to as “passive transport.” Mechanisms requiring applied energy are often called “active transport.”
Active Transport
Cells draw upon metabolic energy to drive solutes across membranes against their concentration gradients. For example, K+ is continuously pumped into human cells as Na+ is pumped out. One source of energy used by these active transporters is the universal cellular currency, adenosine triphosphate (ATP). To tap its energy, plasma membrane-spanning proteins split the ATP to a simpler form (adenosine diphosphate plus phosphate). Instead of allowing the released energy to dissipate as heat, the cleavage step is coupled to the movement of the solute. To accomplish this, the breakdown of the ATP is performed in a pocket of the transporter such that the release of its energy forces the protein into an altered shape. This strain in the protein drives the solute to move “uphill” against its concentration gradient across the membrane, opposite the direction it would spontaneously diffuse. Indeed, the ATP molecules will not be split by the protein unless the solute is being transported simultaneously; the two processes are inextricably coupled through the transporter. A significant fraction of cellular energy is expended in this way.
Membrane Energetics
The conversion of sunlight to electricity by solar panels is a new, “green” alternative to fossil fuels, but nature got there first. Billions of years ago, a membrane transport system evolved that converts solar energy into cellular energy. In this device, photons of sunlight are captured by chlorophyll and other pigments in the membrane. The energy trapped thereby is then used by membrane proteins to force negatively charged electrons away from positively charged protons, separating them across the bilayer. This charge separation turns the membrane into an electrical battery. The potential energy stored in these membranes can then be harvested by proteins that couple it to energy-consuming cellular processes, just as batteries can be used to power a flashlight.
One major use of the potential energy created by separating protons from electrons across membranes is to drive the synthesis of ATP. ATP then powers other metabolic processes, such as the formation of glucose (the major product of photosynthesis), or the transport of solutes discussed in the preceding section. In addition, the membrane potential can itself be directly coupled to pumping certain solutes against their concentration gradient (active transport). In those cases, the downhill diffusion of protons across the membrane provides the energy to pump other solutes uphill. (Picture a paddle wheel that taps a mountain stream for energy to grind grain.) Such membrane potentials also propel bacterial swimming by powering the rotation of their propeller like flagella. Membrane potential energy is thus a currency as universal as ATP or glucose.
Finally, consider how the body’s cells extract energy from glucose. On a gross level, glucose reacts with the oxygen the lungs breathe, yielding energy as well as the carbon dioxide and water the body respires. But this “burning” of glucose is tightly coupled by membrane transport proteins to the pumping of protons and electrons across membranes. Then, just as in photosynthesis, the membrane potential is tapped by other membrane proteins to make ATP or to drive active transport or to enable bacteria to swim.
References
Alberts, Bruce, et al. Molecular Biology of the Cell, 4th ed. New York: Garland Publishing, 2000.
Lodish, Harvey, et al. Molecular Cell Biology, 4th ed. New York: W. H. Freeman and Company, 2000.



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
نحو شراكة وطنية متكاملة.. الأمين العام للعتبة الحسينية يبحث مع وكيل وزارة الخارجية آفاق التعاون المؤسسي
|
|
|