























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Pseudogenes Have Lost Their Original Functions
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 16-3-2021
16-3-2021
 2920
2920




+
-
20
Pseudogenes Have Lost Their Original Functions
KEY CONCEPTS
- Processed pseudogenes result from reverse transcription and integration of mRNA transcripts.
- Nonprocessed pseudogenes result from incomplete duplication or second-copy mutation of functional genes.
- Some pseudogenes might gain functions different from those of their parent genes, such as regulation of gene expression, and take on different names.
pseudogenes are copies of functional genes that have altered or missing regions such that they presumably do not produce polypeptide products with the original function; they can be nonfunctional or have altered function, and the RNA products might serve regulatory functions. For example, as compared to their functional counterparts, many pseudogenes have frameshift or nonsense mutations that disable their protein-coding functionality. There are two types of pseudogenes characterized by their modes of origin.
Processed pseudogenes result from the reverse transcription of mature mRNA transcripts into cDNA copies, followed by their integration into the genome. This might occur at a time when active reverse transcriptase is present in the cell, such as during active retroviral infection or retroposon activity . The transcript has undergone processing , so a processed pseudogene usually lacks the regulatory regions necessary for normal expression. Although it initially contains the coding sequence of a functional polypeptide, it is nonfunctional as soon as it is formed. Such pseudogenes also lack introns and may contain the remnant of the mRNA’s poly(A) tail as well as the flanking direct repeats characteristic of insertion of retroelements .
The second type, nonprocessed pseudogenes, arises from inactivating mutations in one copy of a multiple-copy or single-copy gene or from incomplete duplication of a functional gene. Often, these are formed by mechanisms that result in tandem duplications. An example of a β-globin pseudogene is shown in FIGURE 1. If a gene is duplicated in its entirety with intact regulatory regions, there can be two functional copies for a time, but inactivating mutations in one copy would not necessarily be subject to negative selection. Thus, gene families are ripe for the origin of nonprocessed pseudogenes, as evidenced by the existence of several pseudogenes in the globin gene family . Alternatively, an incomplete duplication of a functional gene, resulting in a copy missing regulatory regions and/or coding sequence, would be “dead on arrival” as an instant pseudogene.
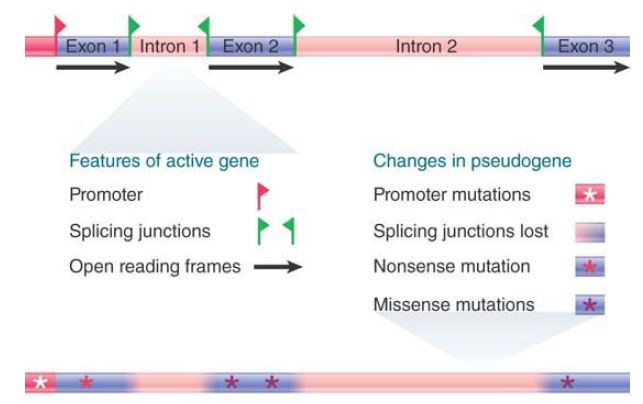
FIGURE 1. Many changes have occurred in a β-globin gene since it became a pseudogene.
There are approximately 20,000 pseudogenes in the human genome. Ribosomal protein (RP) pseudogenes comprise a large family of pseudogenes, with approximately 2,000 copies. These
are processed pseudogenes; presumably the high copy number is a function of the high expression rate of the approximately 80 copies of functional RP genes. Their insertion into the genome is apparently mediated by the L1 retrotransposon . RP genes are highly conserved among species, so it is possible to identify RP pseudogene orthologs in species with a long history of separate evolution and for which whole genome sequences are available.
For example, as shown in TABLE 1, more than two-thirds of human RP pseudogenes are also found in the chimpanzee genome, whereas less than a dozen are shared between humans and rodents. This suggests that most RP pseudogenes are of more recent origin in both primates and rodents, and that most ancestral RP pseudogenes have been lost by deletion or mutational decay beyond recognition.
TABLE 1 Most human RP pseudogenes are of recent origin; many are shared with the chimpanzee but absent from rodents.
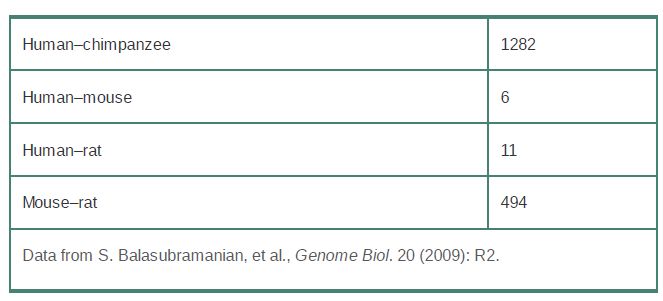
Interestingly, the rate of evolution of RP pseudogenes is slower than that of the neutral rate (as determined by the rate of substitution in ancient repeats across the genome), suggesting negative selection and implying a functional role for RP pseudogenes. Although pseudogenes are nonfunctional when initially formed, there are clear examples of former pseudogenes (originally identified as pseudogenes because of sequence differences with their functional counterparts that would presumably render them nonfunctional) becoming neofunctionalized (taking on a new function) or subfunctionalized (taking on a subfunction or complementary function of the parent gene). When functional again, they would be subject to selection and thus evolve more slowly than expected under a neutral model.
How might a pseudogene gain a new function? One possibility is that translation, but not transcription, of the pseudogene has been disabled. The pseudogene encodes an RNA transcript that is no longer translatable but can affect expression or regulation of the still-functional “parent” gene. In the mouse, the processed pseudogene Makorin1-p1 stabilizes transcripts of the functional Makorin1 gene. Several endogenous siRNAs are encoded by pseudogenes. A second possibility is that a processed pseudogene might be inserted in a location that provides them with new regulatory regions, such as transcription factor binding sites, which allow them to be expressed in a tissuespecific manner unlike that of the parent gene.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















