آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Division Phase of The Cell Cycle |


|
|
Read More
Date: 10-10-2016
Date: 2-11-2016
Date: 9-11-2016
|
Division Phase of The Cell Cycle
The actual division involves two processes: (1) division of the nucleus, called karyokinesis, and (2) division of the protoplasm, called cytokinesis. There are two types of karyokinesis: mitosis and meiosis.
MITOSIS
Mitosis is duplication division, and it is the more common type of karyokinesis. It is the method that any multicellular organism uses as its body is growing and the number of its cells is increasing. It is also used by eukaryotic unicellular organisms when they are not undergoing sexual reproduction. Mitosis is called duplication division because the nuclear genes are first copied; then one set of genes is separated from the other and each is packed into its own nucleus (Fig. 4.11). Each daughter nucleus is basically a duplicate of the original mother nucleus and a twin of the other. Mitosis produces nuclei that are more or less exact copies of the original nucleus, except for occasional errors.
As mentioned above, thousands of genes are linked together into just a few chromosomes, making the separation of gene sets much easier: It is necessary to transport only one of each type of chromosome to each daughter nucleus. This is made even easier because the two new chromatids remain together as the chromosome is replicated in S phase. It is necessary only to make certain that one half of the doubled, large chromosome goes to each end of the cell and the other half to the other end. If that happens with each chromosome, each end of the cell automatically receives one full set of genes. The mechanism that ensures this orderly separation of chromatids is quite straightforward and logical. It consists of the following four phases:
Prophase.
During interphase, the DNA of a chromosome exists as a long, extended double helix associated with histone protein . This open configuration allows enzyme complexes to find specific genes that must be read lor the information they contain. But in this condition, the chromosome may be several centimeters long. It could be wrapped around the cell hundreds of times, making it impossible to pull one chromatid from the other. But during prophase the chromosomes condense that is, they undergo change in the way the histones associate. The chromosomes begin to coil repeatedly, becoming shorter and thicker each time. In addition, a protein framework develops to which the DNA apparently binds. Condensation continues until chromosomes are only 2 to 5 pm long; in this form they can be moved around the cell much more easily.
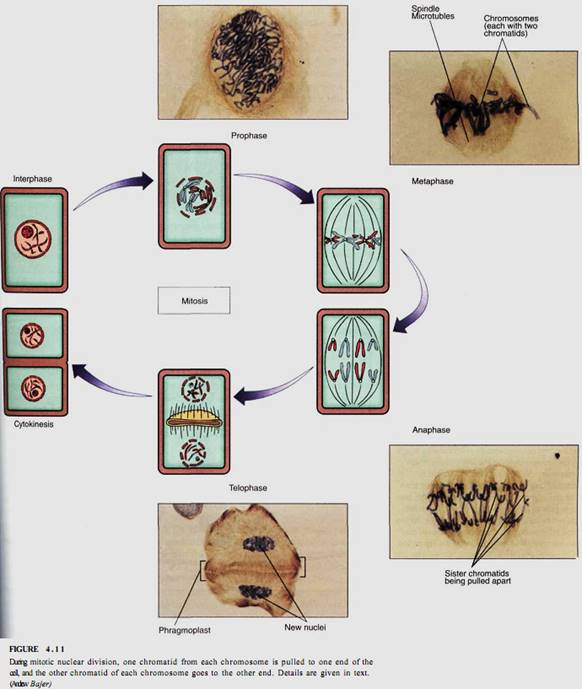
As chromosomes condense and become visible during prophase, other events occur as well (see Fig. 4.11). The nucleolus becomes less distinct and usually completely disappears by the end of prophase. The nuclear envelope also breaks down, apparently owing to the action of enzymes synthesized late in G2. In those organisms that have a set of centrioles (mostly algae, fungi, and animals), the daughter sets that were duplicated during the previous interphase now migrate to opposite poles (sides) of the cell. It had been thought that they simultaneously acted as microtubule-organizing centers and produced a long set of microtubules, the spindle, between themselves. But recent experiments in animals have shown that even if the centrioles are destroyed by laser microbeams, the spindle still forms. It could be that the centrioles themselves are separated by the formation of the spindle, and the association is a mechanism that ensures that each daughter cell receives a centriole.
In the spindle, some microtubules extend from one pole to the center of the cell, where their ends overlap the ends of other microtubules that extend from the opposite end of the spindle. The two sets together, overlapping in the center, form a large framework (Figs. 4.12 and 4.13). Other microtubules run from a pole to a centromere. Each end of the spindle is attached to one of the two faces of the centromere on each chromosome. The point of attachment is a kinetochore; in electron micrographs it appears to be a multilayered structure, but little is known of its actual composition. About 15 to 35 microtubules attach to each kinetochore.


FIGURE 4.12 :(a) Early in mitosis, spindle microtubules attach to the chromosomes at the kinetochore of the centromere. Notice that each chromatid is attached only to the microtubules coming from one end of the cell. (b) Other microtubules pass from pole to pole or overlap the ends of other microtubules at the center of the spindle. As the kinetochore-attached microtubles shorten, the two chromatids of each chromosome are pulled in opposite directions. The long microtubles that do not attach to chromatids act as a framework such that chromatids are pulled apart rather than having the spindle poles pulled into the center of the cell.

Metaphase.
Once the spindle microtubules attach to the centromeres, they push and pull on the chromosomes and gradually move them to the cell center; their arrangement there is called a metaphase plate (see Fig. 4.11). Viewed from the side, the chromosomes appear to be aligned in the very center, but viewed from the end, they are seen to be distributed throughout the central plane of the cell.
No distinct boundary occurs between prophase and metaphase; the chromosomes gradually become visible and gradually move to the metaphase plate. Fragments of nucleolus and nuclear envelope may persist well into metaphase. However, a distinct boundary is evident between metaphase and the following anaphase. At the end of metaphase, the centromeres are duplicated and the two chromatids of each chromosome are suddenly free of each other. In this step, the number of chromosomes is doubled, but the size of each chromosome is halved. Each chromosome is like it was in Gb before S phase. Although the number of chromosomes in the cell is suddenly doubled, the doubling is transitory because the cell soon divides. The exact cause of the separation of the chromatids is not known, but it is thought that the centromere may be a segment of DNA that was not replicated during S phase. Instead, its DNA may be replicated during metaphase and, once it is doubled, the two chromatids are no longer joined.
Anaphase.
Anaphase begins just after the centromere divides; the spindle microtubules that run to the centromeres shorten, depolymerizing at the end near the spindle pole. This is believed to be the primary force responsible for pulling each daughter chromosome away from its twin. Agents that inhibit microtubule depolymerization, such as deuterium oxide, also inhibit chromosome movement, whereas those that speed depolymerization, such as low levels of colchicine, speed movement. The amount of energy necessary to move a chromosome from the metaphase plate to the end of the spindle is small: Just 20 ATP molecules are sufficient. Long chromosomes may tangle somewhat, but microtubules exert sufficient pull to untangle them and drag them to the ends of the spindle. Because the spindle is shaped like a football, as chromosomes on each side get closer to the end, they are pulled together into a compact space.
Telophase.
As chromosomes approach the ends of the spindle, fragments of nuclear envelope appear near them, connect with each other, and form complete nuclear envelopes at each end of the cell. The total surface area of the two new nuclei is larger than that of the envelope of the original mother nucleus; the extra membrane may be derived from endoplasmic reticulum (ER). It is not known how new nuclear pores are formed. Chromosomes become less distinct because they start to uncoil. Gradually, new nucleoli appear as the ribosomal genes become active and produce ribosome subunits. The spindle depolymerizes completely and disappears. Most of the events in telophase are reversals of those in prophase.
To summarize mitosis: After G2 is completed at the end of interphase, each chromosome has been replicated and consists of twin chromatids. Spindle microtubules from opposite poles attach to the centromeres, then pull the twin chromatids of each chromosome away from each other. Two new nuclei form, each containing a full set of chromosomes and each chromosome having one chromatid. The new nuclei are identical to each ether and to the original nucleus that began mitosis. The nuclei can then enter Gi of.
CYTOKINESIS
The division of the protoplast is much simpler than the division of the nucleus. Although it is necessary for each daughter cell to receive some of each type of organelle, random distribution of the organelles in the mother cell usually ensures this. No matter how the cell is divided, each half typically contains some mitochondria, some plastids, some ER, some vacuoles, and so on. It is not necessary for each daughter cell to get exactly half of each type of organelle. A single mitochondrion can divide, or a fragment of ER can grow until the cell has an adequate amount. The same is not true for genes: ff one daughter cell is missing a gene or chromosome, the other genes cannot regenerate the information of the missing gene.
In plants, cytokinesis involves the formation of a phragmoplast made up of short microtubules aligned parallel to the spindle microtubules. The phragmoplast forms in the center of the cell where the metaphase plate had been (Figs. 4.14 and 4.15). Phragmoplast microtubules trap dictyosome vesicles that then fuse into a large, flat, plate like vesicle in which two new primary walls and a middle lamella begin to form. The phragmoplast then grows outward toward the walls of the original cell. It is not known if phragmoplast microtubules actually migrate toward the walls. New microtubules may polymerize near the outer edge of the phragmoplast while those on the inner side depolymerize. New dictyosome vesicles are trapped and added to the edge of the large vesicle; it too grows outward, following the phragmoplast. Similarly, the new walls extend outward along their edges. The phragmoplast, vesicle, and walls are called the cell plate. This process continues until the large vesicle meets the mother cell's plasma membrane, the two fuse, and the vesicle membrane becomes a part of the plasma membranes of the two daughter cells. Simultaneously, the new walls meet and fuse with the wall of the mother cell, completing the division of the mother cell into two daughter cells.

FIGURE 4.14 (a) Late telophase in onion. The row of small vesicles is the beginning of a cell plate. At the edges are short segments of microtubules, the phragmoplast. (b) Cell division has been interrupted by the herbicide DCPA (dimethyl tetrachloroterephalate)

FIGURE 4.15 : (a) Cytokinesis begins as dictyosome vesicles are trapped by phragmoplast microtubules in the space between the two new nuclei, (b) The small vesicles fuse into one large vesicle in which the new middle lamella and two primary walls will form. Plasmodesmata form at this time. (c) The cell plate enlarges toward the existing cell wall as more dictyosome vesicles fuse with the edges of the cell plate vesicle. When the cell plate reaches the existing cell walls, the vesicle membrane fuses with the plasma membrane and thus becomes plasma membrane itself. The new cell plate abuts the old cell wall, and the two become glued together with hemicelluloses and pectins
MEIOSIS
In mitosis, daughter nuclei are replicates of the original mother nucleus. This is necessary for the growth of an organism but creates a problem when sexual reproduction occurs. Two sex cells (gametes) fuse together, forming a zygote, which then grows into a new adult. Each gamete contains one complete set of chromosomes (Fig. 4.16). Nuclei, cells, and
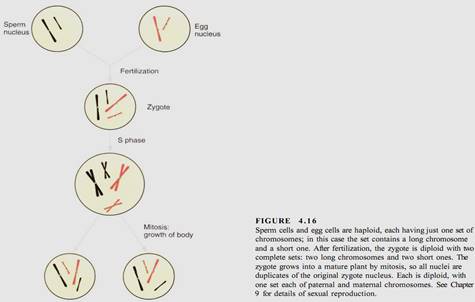
organisms with one set of chromosomes in each nucleus are said to be haploid. The zygote has two complete sets (one from each gamete), so it is diploid. In most species, the zygote grows into an adult by cell divisions in which the nucleus divides by mitosis. As a result, all cells of the adult are diploid because all nuclei are replicates of the mother nucleus.
If the adult were to produce gametes by mitosis, the gametes would be diploid as well, and the next zygote would be tetraploid with four sets. It is not possible biologically to double the number of sets per nucleus with each new generation. It is necessary for a reduction division, called meiosis, to occur somewhere. During reduction division, the two sets of chromosomes present in a diploid nucleus are separated into the daughter nuclei. But a diploid cell is actually faced with separating out four sets of genes because each of the two sets is replicated during the S-phase replication. The cell goes about this in this fashion: Meiosis involves two rounds of division without allowing the S phase to occur after the first division. The two divisions are called meiosis I and meiosis II, and each contains four phases similar to those of mitosis (Fig. 4.17).
Meiosis occurs only in the production of reproductive cells: gametes in animals and some algae and fungi, and spores in plants and other algae and fungi. In seed plants, meiosis occurs only in a few cells in the stamens and ovaries. Meiosis is never used in the growth of the body of any organism.

MEIOSIS I
Prophase I. All the events that characterize prophase of mitosis also occur in prophase I: Nucleolus and nuclear membrane break down; centrioles (if present) separate; a spindle forms; microtubules attach to centromeres; and chromosomes condense and become via-ble (Fig. 4.17). In addition to these processes, special interactions of the chromosomes occur which are unique to prophase I as opposed to prophase of mitosis. Because of this prophase I is divided into five stages:
1. In leptotene, chromosomes begin to condense and become distinguishable, al¬though they appear indistinct.
2. During zygotene, a remarkable pairing of chromosomes occurs (Fig. 4.18a). Re-member that there are two sets of chromosomes, one from the paternal and one from the maternal gamete. Because these have gone through S phase in the preceding interphase, each chromosome has two chromatids; altogether there are four sets of genes on two sets of chromosomes. During zygotene, each chromosome of one set pairs with the equivalent chromosome (its homologous chromosome, or homolog) of the other set; this pairing is synapsis. With remarkable accuracy the two homologous chromosomes in each pair be-come almost perfectly aligned from end to end. A structure, the synaptonemal complex, is present between the paired homologous chromosomes (Fig. 4.18e). This complex is com-posed of a linear central protein element connected by fine transverse fibers to two lateral elements bound to the DNA of the homologous chromosomes.
3. As chromosomes continue to condense, they become shorter and thicker; this stage is pachytene. The synaptonemal complex seems to be involved in the crossing-over that occurs now: In several places in each chromosome, the DNA of each homolog breaks (Fig4.18). Breaks occur in almost identical places in the paired homologs, and the enzymes that repair the breaks hook the "wrong" pieces together. A piece of the maternal homolog is attached to the paternal homolog, and the equivalent piece of the paternal homolog is attached to the maternal one. The full consequences of this are explained in Chapters 16 and 17. If the maternal and paternal chromosomes are absolutely identical, nothing significant has happened. But if the genes on the paternal and maternal chromosomes are slightly different, the new chromosomes that result from synapsis and crossing-over are slightly different from the original chromosomes. There are no new genes, but rather new combinations of genes on each chromatid.
4. After pachytene is diplotene. The homologous chromosomes begin to move away from each other but do not separate completely because they are held together at their paired centromeres and at points (chiasmata; sing.: chiasma) where they appear to be tangled together. Some biologists believe that chiasmata are the points at which crossing- over occurred, but evidence is accumulating that chiasmata are only tangles and are not related to crossing-over at all. Under good conditions it is possible to see all four chromatids of the paired homologous chromosomes; they are called tetrads at this stage.
5. In the final stage, diakinesis, homologs continue to separate, and chiasmata are pushed to the ends of the chromosome. The homologous chromosomes become untangled and are paired only at the centromeres.

Prophase I is the most complicated stage of meiosis; the remaining stages are simple and quite similar to the stages of mitosis.
Metaphase I. Spindle microtubules move the tetrads to the center of the cell, forming a metaphase plate (see Fig. 4.17).
Anaphase I. The homologous chromosomes separate completely from each other, moving to opposite ends of the spindle. The centromeres do not divide, and each chromosome continues to consist of two chromatids. Notice how this is different from the metaphase-anaphase transition of mitosis. In mitosis the centromeres replicate and each chromosome divides into two chromosomes, each with just one chromatid. But in the metaphase I-anaphase I transition, homologous chromosomes separate from each other, and each still has two chromatids. One set of chromosomes is pulled away from the other set, and two new nuclei are formed. These nuclei are now haploid, because each has only one set of chromosomes; the homolog of each chromosome in one nucleus is now in the other nucleus.
Telophase I. Because the chromosomes are still doubled and are in a G2 state, they do not need to undergo an interphase with its G1, S, and G2. Also, because telophase I is basically the opposite of prophase II, some organisms go directly from anaphase I to metaphase II, skipping telophase I and prophase II. In most organisms, however, there is at least a partial telophase I in which chromosomes start to uncoil, and the nucleolus and nuclear envelope start to reappear. If the cells actually progress fully to interphase, no replication of the ENA occurs—the S phase is completely missing. This interval is called interkinesis. Cytokinesis may occur, but it is not unusual for this to be absent also and for both daughter nuclei (i.e., both masses of chromatin) to stay in the original, undivided mother cell..
MEIOSIS II
If a telophase I occurs, then prophase II is necessary to prepare the nucleus for division. Prophase II is not subdivided into stages like prophase I. Metaphase II is short, and at the end of it, the centromeres divide, thereby separating each chromosome into two chiomosomes, just as in metaphase of mitosis, but different from metaphase I. Anaphase II then separates each new chromosome from its replicate, and in telophase II, new nuclei are formed. Each nucleus contains just one set of chromosomes, each with a single chromatid. To summarize, during meiosis I, each chromosome of the paternal chromosomes undergoes synapsis with the homologous chromosome of the maternal set. Crossing-over results in new combinations of genes on the chromosomes. Spindle microtubules pull the paired homologs to opposite poles, so at each pole there is only one set of chromosomes, not two. The new nuclei that form temporarily after meiosis I are thus haploid. When these nuclei undergo meiosis II, the two chromatids of each chromosome are separated, as in mitosis, and the resulting nuclei each have a haploid set of chromosomes, each chromosome with only one chromatid.



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
المقابر الجماعية في العراق.. دراسة في القتل الجماعي والاختفاء القسري بحق الكثير من الأقليات والقوميات العرقية والدينية
|
|
|