آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Bacterial Genetics |


|
Bacterial Genetics
There are hundreds of thousands of bacterial species in existence on Earth. They grow relatively quickly, and most reproduce by binary fission, the production of two identical daughter cells from one mother cell. Therefore, each replication cycle doubles the number of cells in a bacterial population. The bacterial chromosome is a long circle of deoxyribonucleic acid (DNA) that is attached to the membrane of the cell. During replication, the chromosome is copied, and the two copies are divided into the two daughter cells. Transfer of genetic information from the mother cell to offspring is called vertical transmission.
Beneficial mutations that develop in one bacterial cell can also be passed to related bacteria of different lineages through the process of horizontal transmission. There are three main forms of horizontal transmission used to spread genes between members of the same or different species: conjugation (bacteria-to-bacteria transfer), transduction (viral-mediated transfer), and transformation (free DNA transfer). These forms of genetic transfer can move plasmid, bacteriophage, or genomic DNA sequences. A plasmid is a small circle of DNA separate from the chromosome; a bacteriophage is a virus that reproduces in bacteria by injecting its DNA; the genome is the total DNA of the bacterial organism.
After transfer, the DNA molecules can exist in two forms, either as DNA molecules separate from the bacterial chromosome (an episome), or can become part of the bacterial chromosome. The study of basic mechanisms used by bacteria to exchange genes allowed scientists to develop many of the essential tools of modern molecular biology.
A laboratory technician performing an Analytical Profile Index (API) test on bacteria.
Conjugation
Bacterial conjugation refers to the transfer of DNA between bacterial cells that requires cell-to-cell contact. Joshua Lederberg and Edward Tatum firstdescribed conjugation in 1946 when they discovered the F factor (an epi- some) that can move between Escherichia coli cells. The F factor is one of the most well studied conjugative plasmids (plasmids are circular episomes) and is the most well studied conjugative system. There are many different conjugal plasmids carried by members of most bacterial species. Conjugal plasmids that carry antibiotic resistance genes are called R factors. The F factor and R factors usually exist as episomes and each carries functions that allow it to replicate its DNA and thus be inherited by the daughter cells after binary fission. However, conjugative plasmids also express transfer functions that allow the movement of DNA from a donor to a recipient cell; this is the process of conjugation.
The steps of bacterial conjugation are: mating pair formation, conjugal DNA synthesis, DNA transfer, and maturation. The main structure of the F factor that allows mating pair formation is the F pilus or sex pilus (a long thin fiber that extends from the bacterial cell surface). There are one to three pili expressed on an E. coli cell that carries the F factor, and one pilus will specifically interact with several molecules on the recipient cell surface (attachment). About twenty genes on the F factor are required to produce a functional pilus, but the structure is mainly made up of one protein, pilin. To bring the donor and recipient cell into close proximity, the F pilus retracts into the donor cell by removing pilin protein monomers from the base of the pilus to draw the bacterial cells together.
Once a stable mating pair is formed, a specialized form of DNA replication starts. Conjugal DNA synthesis produces a single-stranded copy of the F factor DNA (as opposed to a double-stranded DNA that is formed by normal replication). This DNA strand is transferred into the recipient cell. Once in the recipient cell, the single-stranded copy of the F plasmid DNA is copied to make a double-stranded DNA molecule, which then forms a mature circular plasmid. At the end of conjugation the mating pair is broken and both the donor and the recipient cells carry an identical episomal copy of the F factor. All of the approximately one hundred genes carried on the F factor can now be expressed by the recipient cell and will be inherited by its offspring.
In addition to transferring itself, the F factor can also transfer chromosomal genes between a donor and recipient cell. The F factor can be found inserted (integrated) into the bacterial chromosome at many locations in a small fraction of bacterial cells. An integrated F factor is replicated along with the rest of the chromosome and inherited by offspring along with the rest of the chromosome. When a mating pair is formed between the donor cell carrying an integrated F factor and a recipient cell, DNA transfer occurs as it does for the episomal F factor, but now the chromosomal sequences adjacent to the integrated F factor are transferred into the recipient. Since these DNA sequences encode bacterial genes, they can recombine with the same genes in the recipient. If the donor gene has minor changes in DNA sequence from the recipient gene, the different sequence can be incorporated into the recipient gene and inherited by the recipient cell’s offspring. Donor cells that have an integrated copy of the F factor are called Hfr strains (High frequency of recombination).
Transduction
The second way that DNA is transferred between bacterial cells is through a phage particle in the process of transduction. Joshua Lederberg and Norton Zinder first discovered transduction in 1956. When phage inject their DNA into a recipient cell, a process occurs that produces new bacteriophage particles and kills the host cell (lytic growth). Some phage do not always kill the host cell (temperate phage), but instead can be inherited by daughter host cells. Therefore acquisition of a so-called temperate “prophage” by a recipient cell is a form of transduction. Many phage also have the ability to transfer chromosomal or plasmid genes between bacterial cells. During generalized transduction any gene can be transferred from a donor cell to a recipient cell. Generalized transducing phages are produced when a phage packages bacterial genes into its capsid (protein envelope) instead of its own DNA. When a phage particle carrying bacterial chromosomal genes attaches to a recipient cell, the DNA is injected into the cytoplasm where it can recombine with a homologous DNA sequences.
Some bacteriophage can pick up a subset of chromosomal genes and transfer them to other bacteria. This process is called specialized transduction since only a limited set of chromosomal genes can be transferred between bacterial cells.
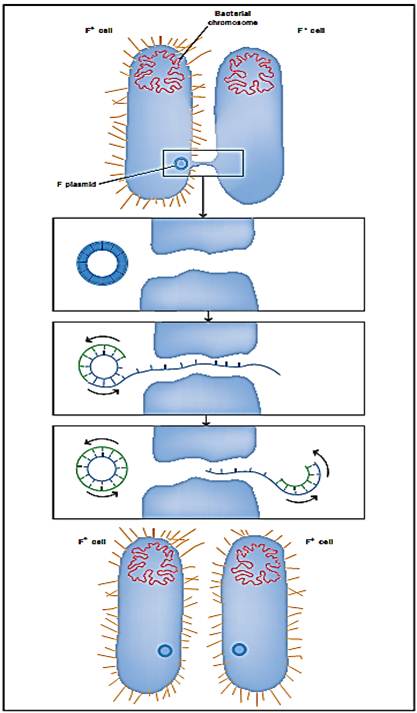
Bacterial conjugation. The bacterium on the left passes a copy of the F plasmid to the bacterium on the right, converting it from an F“ cell to an F+ cell.
Transformation
The third main way that bacteria exchange DNA is called DNA transformation. Some bacteria have evolved systems that transport free DNA from the outside of the bacterial cell into the cytoplasm. These bacterial are called “naturally competent” for DNA transformation. Natural DNA transformation of Streptococcus pneumonaiae provided the first proof that DNA encoded the genetic material in experiments by Oswald Avery and colleagues. Some other naturally competent bacteria include Bacillus subtilis, Haemophilus influenzae, and Neisseria gonorrhoeae. Other bacterial species such as E. coli are not naturally competent for DNA transformation. Scientists have devised many ways to physically or chemically force noncompetant bacteria to take up DNA. These methods of artificial DNA transformation form the basis of plasmid cloning in molecular biology.
Most naturally competent bacteria regulate transformation competence so that they only take up DNA into their cells when there is a high density of cells in the environment. The ability to sense how many other cells are in an area is called quorum sensing. Bacteria that are naturally competent for DNA transformation express ten to twenty proteins that form a structure that spans the bacterial cell envelope. In some bacteria this structure also is required to form a particular type of pilus different than the F factor pilus. Other bacteria express similar structures that are involved in secreting proteins into the exterior medium (Type II secretion). Therefore, it appears that DNA transformation and protein secretion have evolved together.
During natural DNA transformation, doubled-stranded DNA is bound to the recipient cell surface by a protein receptor. One strand of the DNA is transported through the cell envelope, where it can recombine with similar sequences present in the recipient cell. If the DNA taken up is not homologous to genes already present in the cell, the DNA is usually broken down and the nucleotides released are used to synthesize new DNA during normal replication. This observation has led to the speculation that DNA transformation competence may have originally evolved to allow the acquisition of nucleic acids for food.
The source of DNA for transformation is thought to be DNA released from other cells in the same population. Most naturally competent bacteria spontaneously break apart by expressing enzymes that break the cell wall. Autolysis will release the genomic DNA into the environment where it will be available for DNA transformation. Of course, this results in the death of some cells in the population, but usually not large numbers of cells. It appears that losing a few cells from the population is counterbalanced by having the possibility of gaining new traits by DNA transformation.
References
Tortora, Gerard J., Berdell R. Funke, Christine L. Case. Microbiology: An Introduction. Redwood City: CA: Benjamin/Cummings Publishing Company, Inc., 2001.



|
|
|
|
مخاطر خفية لمكون شائع في مشروبات الطاقة والمكملات الغذائية
|
|
|
|
|
|
|
"آبل" تشغّل نظامها الجديد للذكاء الاصطناعي على أجهزتها
|
|
|
|
|
|
|
المجمع العلميّ يُواصل عقد جلسات تعليميّة في فنون الإقراء لطلبة العلوم الدينيّة في النجف الأشرف
|
|
|